ความอร่อยและความสมดุลของอาหารเป็นปัจจัยที่สัมพันธ์กัน วิธีการศึกษาฟังก์ชันรสชาติ
เป็นต้นฉบับ
4 LEK PP" Nikolaeva Ekaterina Valerievna
ศึกษาความชอบด้านรสชาติของปลา
มอสโก - 2000
งานนี้ดำเนินการที่สถาบันนิเวศวิทยาและวิวัฒนาการ A.N. Severtsov ของ Russian Academy of Sciences และที่คณะชีววิทยาของ M.V. Lomonosov Moscow State University
ที่ปรึกษาทางวิทยาศาสตร์:
ที่ปรึกษาด้านวิทยาศาสตร์:
นักวิชาการของ Russian Academy of Sciences D.S. Pavlov
ศาสตราจารย์ ดร. วิทยาศาสตรบัณฑิต ชีววิทยา อ.กสุมญาณ
คู่ต่อสู้อย่างเป็นทางการ: ศาสตราจารย์, หมอ
วิทยาศาสตร์ชีวภาพ Yu.B. Manteifel
ผู้สมัคร
วิทยาศาสตร์ชีวภาพ V.M.Stygar
องค์กรชั้นนำ: มอสโกการเกษตร
Academy ตั้งชื่อตาม K.A. Timiryazev
การป้องกันวิทยานิพนธ์จะมีขึ้นในวันที่ 19 ธันวาคม 2543 เวลา 10.00 น. ในการประชุมสภาวิทยานิพนธ์ D 002.48.01 เพื่อการป้องกันวิทยานิพนธ์ในระดับผู้สมัครสาขาวิทยาศาสตร์ชีวภาพที่ A.N. Severtsov Institute of Ecology และ วิวัฒนาการ RAS, 117071 มอสโก, Leninsky Prospekt, 33.
วิทยานิพนธ์สามารถพบได้ในห้องสมุดของภาควิชาชีววิทยาทั่วไปของ Russian Academy of Sciences
เลขาธิการสภาวิทยาศาสตร์ L.T. Kapralova
: ผู้สมัครสาขาวิทยาศาสตร์ชีวภาพ
ปอนด์ซ. M - /¿A Sh ok ML 3 . ป
ความเกี่ยวข้องของหัวข้อ การรับรู้รสชาติมีบทบาทสำคัญในการสนับสนุนทางประสาทสัมผัสในช่วงสุดท้ายของพฤติกรรมการกินอาหารที่เกี่ยวข้องกับการประเมินคุณสมบัติด้านรสชาติของเหยื่อและการปฏิบัติตามความต้องการทางโภชนาการของปลา และการใช้รายการอาหารที่เพียงพอ (Atema, 1980;
พาฟโลฟ, กะสุมยาน, 2533) การศึกษาเกี่ยวกับระบบการรับรสของปลาจำนวนมากที่ดำเนินการจนถึงปัจจุบัน มุ่งเน้นไปที่การชี้แจงลักษณะทางสัณฐานวิทยาและภูมิประเทศของปุ่มรับรส โครงสร้างพิเศษ และการปกคลุมด้วยเส้นของมันเป็นหลัก ศึกษาการจัดโครงสร้างทางสัณฐานวิทยาของส่วนกลาง (Reutter, 1986; Jakubowski, Whiter, 1990) ศึกษา คุณสมบัติการทำงานการศึกษาระบบการรับรสจะดำเนินการในกรณีส่วนใหญ่โดยใช้วิธีอิเล็กโทรสรีรวิทยา (Marui, Caprio, 1992) วิธีการประเมินความชอบด้านรสชาติของปลายังคงพัฒนาได้ไม่ดีและเป็นผลให้ข้อมูลเกี่ยวกับทัศนคติของปลาต่อรสชาติของสารประเภทต่างๆ แทบไม่มีเลยจนกระทั่งเมื่อไม่นานมานี้ ปริมาณข้อมูลการทดลองที่ได้รับโดยใช้ปฏิกิริยาทดสอบพฤติกรรมและเผยให้เห็นถึงรสชาติที่ชอบของปลานั้นมีจำกัดและไม่ได้ให้แนวคิดที่ชัดเจนเกี่ยวกับรูปแบบทั่วไปและลักษณะเฉพาะของทัศนคติของปลาต่อรสชาติสิ่งเร้า ความเหมือนและความแตกต่างใน สเปกตรัมของสารปรุงแต่งรสที่มีประสิทธิผลในปลาที่มีรูปแบบการใช้ชีวิตที่แตกต่างกันและการจัดเตรียมที่เป็นระบบ (Kasumyan, 1997)
อิทธิพลต่อความไวต่อรสชาติของปลาของปัจจัยทางชีวภาพและปัจจัยทางชีวภาพต่างๆ สภาพความเป็นอยู่ของปลา และลักษณะของโภชนาการของปลายังคงมีการศึกษาที่ไม่ดีนัก คำถามเกี่ยวกับการมีอยู่และธรรมชาติของการเชื่อมโยงระหว่างความชอบด้านรสชาติกับความกว้างของสเปกตรัมอาหารของปลา องค์ประกอบของสิ่งมีชีวิตในอาหารที่บริโภค และประสบการณ์ทางโภชนาการของแต่ละบุคคลยังคงเปิดกว้างอย่างสมบูรณ์ ไม่มีข้อมูลเกี่ยวกับความรุนแรงของพฟิสซึ่มทางเพศในรสชาติของปลา
การตั้งค่าเกี่ยวกับอิทธิพลของวิถีชีวิตของปลาที่มีต่อการสำแดงองค์ประกอบต่าง ๆ ของการตอบสนองต่อพฤติกรรมต่อสิ่งเร้ารสชาติ
กำลังศึกษาสิ่งเหล่านี้และอื่นๆ ปัญหาปัจจุบันการรับรู้รสชาติของปลามีความสำคัญทางทฤษฎีที่สำคัญ เนื่องจากทำให้สามารถชี้แจงบทบาทของระบบประสาทสัมผัสในการเลือกวัตถุอาหารที่เพียงพอของปลาได้ ความรู้เรื่องรูปแบบ ความไวต่อรสชาติปลาคุณสมบัติเฉพาะของการตอบสนองต่อสารปรุงแต่งชนิดต่าง ๆ นั้นเป็นที่สนใจในทางปฏิบัติอย่างไม่ต้องสงสัยและสามารถนำมาใช้ในการแก้ปัญหาต่าง ๆ ในการประมงและการเพาะเลี้ยงสัตว์น้ำ วัตถุประสงค์และวัตถุประสงค์ของการศึกษา งานวิจัยนี้มีวัตถุประสงค์เพื่อศึกษาลักษณะความชอบด้านรสชาติของปลา งานของงานประกอบด้วย:
การเปรียบเทียบความชอบด้านรสชาติของปลาที่อาศัยอยู่ในน้ำที่มีความเค็มต่างกัน
การวิจัยเกี่ยวกับความสัมพันธ์ระหว่างรูปแบบการให้อาหาร สเปกตรัมอาหาร และความชอบด้านรสชาติของปลา
ศึกษาอิทธิพลของประสบการณ์ด้านอาหารต่อความชอบด้านรสชาติของปลา
การกำหนดลักษณะการสืบทอดความชอบด้านรสชาติของปลา
การศึกษาความหลากหลายทางเพศของรสนิยมในปลา (โดยใช้ตัวอย่างของปลาหางนกยูง)
ชี้แจงคุณลักษณะของการสำแดงองค์ประกอบหลักของการตอบสนองพฤติกรรมการรับรสของปลาในระบบนิเวศที่แตกต่างกัน
ความแปลกใหม่ทางวิทยาศาสตร์ ความแปลกใหม่ทางวิทยาศาสตร์ของวิทยานิพนธ์นี้อยู่ที่การขยายความเข้าใจเกี่ยวกับลักษณะการทำงานของระบบรสชาติของปลา ผลงานเผยเป็นครั้งแรกถึงรสชาติของปลา 7 สายพันธุ์สำหรับสารปรุงแต่งรสคลาสสิก และปลา 4 สายพันธุ์สำหรับกรดอะมิโนอิสระ เป็นครั้งแรกที่มีการกำหนดจุดยืนเกี่ยวกับการไม่มีความสัมพันธ์ระหว่างความชอบด้านรสชาติของปลากับพวกมัน
ความสัมพันธ์กับความเค็มของน้ำ ระหว่างระดับยูริฟาจีของปลากับความกว้างของสเปกตรัมของสารกระตุ้น พบว่าปลาที่มีส่วนประกอบจากพืชชัดเจนมีทัศนคติเชิงบวกต่อน้ำตาล (ซูโครส)
เป็นครั้งแรกสำหรับบุคคลที่มีไอโซเจนิกที่เติบโตบน หลากหลายชนิดอาหาร แสดงให้เห็นว่าความชอบด้านรสชาติของปลาไม่ได้ถูกแก้ไขโดยประสบการณ์ด้านอาหาร อยู่ภายใต้การควบคุมทางพันธุกรรมอย่างเข้มงวดและการถ่ายทอดทางพันธุกรรมของพวกมันเป็นไปตามธรรมชาติของ Patroclinic การใช้ตัวอย่างของปลาหางนกยูงที่โตเต็มวัยทางเพศ การขาดความหลากหลายทางเพศในการตั้งค่ารสชาติของปลา ได้รับการจัดตั้งขึ้นแสดงให้เห็นว่าความแตกต่างทางเพศสามารถแสดงออกมาอย่างรุนแรงโดยปลาของพารามิเตอร์จำนวนหนึ่งของการตอบสนองต่อรสชาติเชิงพฤติกรรม พบว่าในปลาที่มีวิถีชีวิตที่แตกต่างกันการแสดงองค์ประกอบบางอย่างของการตอบสนองรสชาติเชิงพฤติกรรม แตกต่างกัน (จำนวนการจับเม็ด ระยะเวลาการยึดเม็ดเข้า) ช่องปาก). เป็นที่ยอมรับแล้วว่าประสบการณ์การรับประทานอาหารที่แตกต่างกันไม่ส่งผลกระทบต่อรูปแบบการตอบสนองรสชาติโดยรวมของปลา
ความสำคัญในทางปฏิบัติ ผลการศึกษานี้สามารถใช้เป็นพื้นฐานในการพัฒนาและแนะนำการเพาะเลี้ยงสัตว์น้ำด้วยสารเคมีกระตุ้นรสชาติที่ออกแบบมาเพื่อเพิ่มประสิทธิภาพของสูตรอาหารเทียมเพื่อเพิ่มรสชาติให้น่ารับประทานสำหรับปลาในฟาร์ม ผลลัพธ์ที่ได้สามารถนำไปใช้ในอุตสาหกรรมตู้ปลาได้ (ปลาหางนกยูง ปลาหมอสี) และกีฬาตกปลา (แมลงสาบ ปลาคาร์พเงิน)
ผลการศึกษานี้ใช้เป็นส่วนหนึ่งของหลักสูตรการบรรยาย "สรีรวิทยาของปลา" ซึ่งมอบให้กับนักศึกษาภาควิชา Ichthyology คณะชีววิทยา มหาวิทยาลัยแห่งรัฐมอสโก
การอนุมัติงาน วิทยานิพนธ์ถูกนำเสนอในการประชุมนานาชาติเรื่อง Ichthyofauna ของภาคเหนือ (เบอร์เกน, นอร์เวย์, 1993) การประชุม
“ปัญหาการเรียน การใช้เหตุผลและการคุ้มครองทรัพยากรธรรมชาติของทะเลสีขาว" (เซนต์ปีเตอร์สเบิร์ก, 1995), การประชุม All-Russian ครั้งที่ 2 เรื่องพฤติกรรมปลา (Borok, 1996), การประชุม XXVII ของนักวิทยาศาสตร์รุ่นเยาว์ของคณะชีววิทยาของมหาวิทยาลัยแห่งรัฐมอสโก (มอสโก, 1997) การประชุมเชิงปฏิบัติการของห้องปฏิบัติการพฤติกรรมของสัตว์มีกระดูกสันหลังส่วนล่างของสถาบันปัญหาระบบนิเวศและวิวัฒนาการของนาโนเมตร A.N. Severtsov RAS และห้องปฏิบัติการเคมีบำบัดและเคมีสื่อสารของปลาของภาควิชา Ichthyology แห่งมหาวิทยาลัยแห่งรัฐมอสโก
สิ่งพิมพ์ บทบัญญัติหลักของวิทยานิพนธ์แสดงไว้ในงานพิมพ์ 6 เรื่อง
โครงสร้างและขอบเขตของงาน วิทยานิพนธ์ประกอบด้วยคำนำ 4 บท บทสรุป และรายการอ้างอิง ต้นฉบับประกอบด้วย
หน้าซึ่ง.....หน้าข้อความพิมพ์ดีด 7 รูปวาด
และ 33 โต๊ะ บรรณานุกรมประกอบด้วยผลงานในประเทศ 133 ชิ้น และนักเขียนชาวต่างประเทศ 151 ชิ้น
วัสดุและวิธีการ
งานทดลองได้ดำเนินการที่ภาควิชา Ichthyology ของคณะชีววิทยาและสถานีชีววิทยาทะเลสีขาวของมหาวิทยาลัยแห่งรัฐมอสโกตลอดจนในห้องปฏิบัติการพฤติกรรมของสัตว์มีกระดูกสันหลังส่วนล่างของสถาบันเศรษฐกิจเชิงนิเวศน์ที่ตั้งชื่อตาม A.N. Severtsov RAS ในช่วงปี 1989 ถึง 1998 วัตถุประสงค์ของการศึกษาครั้งนี้คือปลา 7 ชนิด:
Roach Rutilus rutilus อายุสองปี ยาว (L) 6.5 ซม.
ปลาทอง Carassius auratus gibelio, ลูกปลา, ความยาว (L) 6.5 ซม.;
Guppies Poeciia reticulata อายุ 6-8 เดือน ความยาว (L) 2.5 ซม.
Cichlasoma severum Heros (Cichlasoma) severus อายุ 4-5 เดือน ความยาว (L) b cm;
ปลาปักเป้าหลังหนามเก้าหนาม Pungitius pungitius อายุปี ความยาว (L) 4.8 ซม.
ปลาลิ้นหมาอาร์กติก Liopsetta glacialis อายุ 6 เดือน ความยาว (L) 6.3 ซม.
ปลาดุกลาย Anarhichas lupus อายุ 5-6 เดือน ความยาว (L)
สำหรับงานทดลอง เรายังใช้บุคคลทางจีโนเจเนติกส์ (อายุปี 4-5 ซม.) ของลูกผสมระหว่างปลาคาร์พสีเงิน crucian (ตัวเมีย) และปลาคาร์พ Cyprinus carpió (ตัวผู้) ที่ได้รับจาก VNPO สำหรับการเลี้ยงปลา (Dmitrov ภูมิภาคมอสโก)
ปลาจะเคยชินกับสภาพการทดลองตั้งแต่ 2 สัปดาห์ถึงหลายเดือน ในเวลานี้ พวกเขาถูกเก็บไว้ในตู้ปลาทั่วไปในอุณหภูมิที่เหมาะสมที่สุดสำหรับแต่ละสายพันธุ์ และเลี้ยงด้วยหนอนเลือดที่มีชีวิตหรือแช่แข็งสด (ลูกน้ำยุงลาย Chironomidae) หรือเนื้อปลา (สายพันธุ์ทะเล) การทดลองเพื่อกำหนดอิทธิพลของประสบการณ์ด้านอาหารต่อความชอบด้านรสชาติของปลาได้ดำเนินการกับปลาลูกผสมปลาคาร์พ crucian ลูกผสม (ลูกนิ้ว) หลังจากจับจากบ่อได้แบ่งออกเป็น 3 กลุ่ม และอีก 6 เดือนต่อมาก็ถูกเลี้ยงในตู้ปลาต่างๆ และเลี้ยงด้วยอาหารเทียมสำหรับ ตู้ปลา“Tetramin” (ผลิตโดย Tetra ประเทศเยอรมนี กลุ่มหมายเลข 1) tubifex ที่มีชีวิต (oligochaetes ของสกุล Tubifex กลุ่มหมายเลข 2) และหนอนเลือดสดหรือแช่แข็งสด (กลุ่มหมายเลข 3)
1-2 สัปดาห์ก่อนเริ่มการทดลอง ปลาจะถูกวางทีละตัวในตู้ปลาขนาดเล็ก (5-12 ลิตร) ผนังด้านหลังและด้านข้างของตู้ปลาทำจากพลาสติกไวนิลทึบแสงเพื่อแยกปลาออกจากกัน การสังเกตปลาได้ดำเนินการผ่านผนังด้านหน้าโปร่งใส การทดลองกับปลาลิ้นหมาขั้วโลกและปลาดุกลายได้ดำเนินการในการตั้งค่าการทดลองซึ่งประกอบด้วยช่องประเภทการไหลที่เชื่อมต่อถึงกัน (ช่องละ 25 ลิตร) โดยมีการป้อนปลาฟลาวเดอร์อย่างต่อเนื่อง น้ำทะเล. เพื่อประเมินความชอบด้านรสชาติของปลาต่อสิ่งเร้าทางเคมีจึงใช้วิธีที่เสนอโดย A.O. Kasumyan และ S.S. Sidorov (Kasumyan, Sidorov, 1992,1993) ประกอบด้วยการบันทึกพฤติกรรมการตอบสนองของปลาตัวเดียว
เม็ดเจลวุ้นวุ้นเทียมที่มีสารทดสอบอย่างใดอย่างหนึ่ง สารเคมี. ในงานนี้ กรดอะมิโนอิสระ (บี-สเตอรีโอไอโซเมอร์) และสารแต่งกลิ่นรสดั้งเดิมที่ทำให้เกิดสารประเภทหลัก ลิ้มรสความรู้สึกในมนุษย์ สารที่มีความบริสุทธิ์ทางเคมีในระดับสูงถูกนำมาใช้เป็นสารระคายเคือง รายชื่อสารแต่งกลิ่นที่ใช้และความเข้มข้นแสดงไว้ในตารางที่ 1
ก่อนการทดลองเริ่มต้น สัตว์ทดลองได้รับการฝึกให้จับเม็ดที่มีสารสกัดจากอาหาร (75 กรัม/ลิตร) ในระหว่างการทดลอง มีการบันทึกสิ่งต่อไปนี้: 1) ความสามารถในการรับประทานของเม็ด ได้แก่ เม็ดที่ถูกจับถูกกินหรือถูกปฏิเสธ 2) จำนวนการกระทำของการตั้งค่าเม็ดเล็ก 3) ระยะเวลาในการคงตัวของเม็ดหลังจากการตั้งค่าครั้งแรก 4) ระยะเวลารวมของการเก็บรักษาแกรนูลระหว่างการทดลองทั้งหมด เริ่มลงทะเบียนตั้งแต่วินาทีที่เม็ดชุดแรก ช่วงเวลาที่ปลากลืนเม็ดจะถูกกำหนดโดยการเคลื่อนไหวเคี้ยวของกรามในลักษณะเฉพาะและการฟื้นฟูการเคลื่อนไหวเป็นจังหวะโดยฝาครอบเหงือก ระยะเวลาที่ปลากักเศษเม็ดในช่องปากไว้ถูกบันทึกโดยใช้นาฬิกาจับเวลาแบบสรุปของ Agat ปลาจะสุ่มให้เม็ดที่มีสารต่างกันตามลำดับ การจัดหาเม็ดที่มีสารทดสอบอย่างใดอย่างหนึ่งสลับกับการจัดหาเม็ดที่มีสารสกัดอาหารสัตว์ ในการทดลองควบคุม จะใช้แกรนูลที่มีเพียงสีย้อมเท่านั้น การทดลองที่เม็ดไม่สามารถจับได้ภายใน 1 นาทีนับจากวินาทีที่นำมาให้ปลาไม่ได้ถูกนำมาพิจารณา ในการทดลองกับปลาดุกลายนั้น ปลาทดลองจะถูกป้อนพร้อมกัน 10 แผ่นวุ้น-วุ้น (4x4x2 มม.) ที่มีสารปรุงแต่งรสที่ผ่านการทดสอบแล้ว หลังจากผ่านไปหนึ่งนาที ก็นับจำนวนปลาที่ยังไม่ได้กิน
จานซึ่งถูกเอาออกจากตู้ปลาแล้ว ช่วงเวลาระหว่างการทดลองแต่ละครั้งกับผู้ทดลองคนเดียวกันในชุดการทดลองทั้งหมดคืออย่างน้อย 10-15 นาที
เตรียมแกรนูลทันทีก่อนเริ่มการทดลองจากเจลวุ้นวุ้น (Reanal 2%) โดยเติมสารทดสอบตัวใดตัวหนึ่งหรือสารสกัดอาหารสัตว์ระหว่างการเตรียม เพื่อให้เม็ดเห็นได้ชัดเจนยิ่งขึ้นในเนื้อเจล เติมสีย้อมเพื่อให้ได้สีแดงสด (ปอนโซ 4R, 0.0005 M) หรือ สีเขียว(Cr203.0.3% - ในการทดลองกับปลาหมอสี cichlasoma severum และปลาดุกลาย) ขนาดของเม็ดสอดคล้องกับขนาดของปลาทดลองและสามารถกลืนได้ง่าย สำหรับแมลงสาบ ปลาหางนกยูง และสติ๊กเกิลแบ็กเก้าหนาม เส้นผ่านศูนย์กลางของเม็ดคือ 1.5 มม. โดยมีความยาวเม็ด 2.5 มม. สำหรับปลาคาร์พ crucian เงิน ปลาลิ้นหมาอาร์กติก และลูกผสมของปลาคาร์ป crucian เส้นผ่านศูนย์กลางของเม็ดคือ 2.0 มม. โดยมีความยาวเม็ด 3.0 มม. เจลที่มีกรดอะมิโนหรือสารแต่งกลิ่นรสคลาสสิกเก็บไว้ที่อุณหภูมิ +5° C ไม่เกิน 7 วัน เจลที่มีสารสกัดอาหารจะถูกเก็บไว้ภายใต้สภาวะเดียวกันไม่เกิน 3-4 วัน
การประมวลผลวัสดุหลักดำเนินการโดยใช้ชุดซอฟต์แวร์ Statgraphics (เวอร์ชัน 3.0), MatCad และ Microsoft Excel (7.0)97 การวิเคราะห์ทางสถิติผลลัพธ์ดำเนินการโดยใช้การทดสอบ I ของนักเรียนและสัมประสิทธิ์สหสัมพันธ์อันดับของสเปียร์แมน (r5) ทำการทดลองทั้งหมด 17,372 ครั้ง
1. รสชาติตอบสนองต่อสารรสคลาสสิกและ
กรดอะมิโนอิสระในปลาที่มีทัศนคติต่อความเค็มของน้ำและรูปแบบการให้อาหารที่แตกต่างกัน
วัตถุประสงค์ประการหนึ่งของงานนี้คือเพื่อชี้แจงลักษณะความชอบด้านรสชาติของปลาที่อาศัยอยู่ในน้ำที่มีความเค็มต่างกัน การวิจัยได้ดำเนินการ
ปลาน้ำจืด - แมลงสาบ, ปลาคาร์พสีเงิน crucian, ปลาหางนกยูงและปลาซิคลาโซมาเซเวอร์รัม, สายพันธุ์ยูริฮาลีน - ปลาสลิดเก้าหนามและปลาทะเล - ปลาลิ้นหมาอาร์กติกและปลาดุกลาย ดอกเบี้ยมากที่สุดนำเสนอการเปรียบเทียบพันธุ์ปลาตามรายการรสชาติที่ตอบสนองต่อโซเดียมคลอไรด์และแคลเซียมคลอไรด์ ซึ่งเป็นสารที่ ส่วนประกอบที่สำคัญน้ำธรรมชาติ การทดลองแสดงให้เห็นว่าสำหรับแมลงสาบและปลาลิ้นหมาอาร์กติก โซเดียมคลอไรด์เป็นตัวกระตุ้นรสชาติที่เพิ่มการบริโภคอาหารเม็ด สำหรับปลาคาร์พ crucian เงินและปลาหมอสี เป็นตัวยับยั้งที่ช่วยลดการบริโภคอาหารเม็ดได้อย่างมาก ปลาหางนกยูง ปลาสติ๊กเกิลแบ็กเก้าหนาม และปลาดุกลายมีทัศนคติที่ไม่แยแสต่อเม็ดเล็กๆ ที่มีสารนี้ แคลเซียมคลอไรด์เป็นสารกระตุ้นรสชาติเฉพาะสำหรับปลาซิคลาโซมา เซเวอร์รัม สำหรับน้ำจืดอื่นๆ และ ปลาทะเลเราศึกษามันทำหน้าที่เป็นสารรับรสที่ไม่แยแส (ตารางที่ 1) ข้อมูลจากวรรณกรรมยืนยันตำแหน่งนี้ ปลาน้ำจืดหรือปลาทะเลที่ศึกษาโดยผู้เขียนคนอื่นๆ มักแสดงการตอบสนองต่อรสชาติที่แตกต่างกันหรือตรงกันข้ามกับสิ่งเร้ารสชาติเดียวกัน และมักน้อยกว่ามากด้วยซ้ำกับการตอบสนองประเภทเดียวกัน ปลาน้ำจืด เช่น dace Leuciscus leuciscus, ปลาคาร์พหญ้า Ctenopharyngodon idella, crimson loach Salvelinus namaycush, European greyling Thymallus thymallus และ Anadromous Caspian brown trout Salmo trutta caspius แสดงถึงรสนิยมของโซเดียมคลอไรด์ ในปลาสเตอร์เจียนไซบีเรียน้ำจืด Acipenser baerii, ปลาสเตอร์เจียน stellate ชนิด Anadromous Acipenser stellatus และ หญ้าฝรั่นทะเลจากกรณีนี้ การรวมโซเดียมคลอไรด์ไว้ในเม็ดทำให้การบริโภคเม็ดลดลงอย่างมีนัยสำคัญ (Kasumyan, Sidorov, 1993; Kasumyan et al., 1993a; Kasumyan, Kazhlaev, 1993; Kasumyan, Sidorov, 1995a; Kasumyan, 1997; คาสุมยาน, โมเรย์, 1997) จากแหล่งวรรณกรรมเดียวกันเป็นที่ทราบกันว่าแคลเซียมคลอไรด์ทำให้การบริโภคเพิ่มขึ้น
ตารางที่ 1. รสชาติดึงดูดใจสารปรุงแต่งรสดั้งเดิมและกรดอะมิโนอิสระในปลาที่อาศัยอยู่ในน้ำที่มีความเค็มต่างกัน
("+" - ผลกระตุ้น, "-" - ผลการยับยั้ง, 0 - คุณสมบัติรสชาติที่ไม่แยแส)
p/n ระคายเคือง Concentration, M Roach ปลาคาร์ปสีเงิน crucian Guppy Cichlasoma severum ปลาลิ้นหมาหลังหนามเก้าหนาม ปลาลิ้นหมาอาร์กติก ปลาดุกลาย
1 โซเดียมคลอไรด์ 1.73(10%) ttya - 0 - 0 0
2 แคลเซียมคลอไรด์ 0.9(10%) 0 0 0 0 0 0
3 กรดซิตริก 0.26 (5%) - - .
4 ซูโครส 0.29(10%) 0 จำเป็น! 0 0 0 0
5 ไกลซีน 0.1 0 0
6 อลันย่า 0.1 0 0 0
7 เซริน 0.1 vzhsha shashsh 0 0
8 โพรลีน 0.1 0 . 0 0
9 ซิสเทอีน 0.1 0 0 0
10 กรดกลูตามิก 0.01 0 0 0
11 กลูตามีน 0.1 จุด 0 Ш8ШЯ 0
12 กรดแอสลาราจิก 0.01 0 0 0 0
13 แอสพาราจีน 0.1 0 0 0 0
14 ทรีโอนีน 0.1 ¡ SHED 0 0
15 ทริปโตเฟน 0.01 0 0 0
16 ไทโรซีน 0.001 เมียด-ย่า 0 0
17 นอร์วาลีน 0.1 อาดายาซ 0 0 0
18 วาลิน 0.1 0 EEZZKZZZZ 0 0
19 ฟีนิลอะลานีน 0.1 0 - 0 0
20 เมไทโอนีน 0.1 0 0 0 0
21 ลิวซีน 0.01 0 shsh+tt 0 0
22 ไลซีน 0.1 0 0 886*355$ 0
23 ไอโซลิวซีน 0.01 0 0 0
24 อาร์จินีน 0.1 VZh+ZhZ 0 0
25 ฮิสติดีน 0.1 0 0 0 0
เม็ดในปลาคาร์พ ปลา chub leiodcus serballus ปลา cristivomer char ปลาเกรย์ยุโรป ปลาเรนโบว์เทราต์ Parabalico mysus และปลาเทราท์สีน้ำตาลแคสเปียน และปลาสเตอร์เจียนไซบีเรียทำให้การบริโภคปลาเม็ดลดลง (Kasumyan, Sidorov, 1993; Kasumyan, Kazhlaev, 1993; Kaetuan, 51(Iogou, 1995a; Kasumyan, Morey, 1996; Kasumyan, 1997)
การเปรียบเทียบความชอบด้านรสชาติของปลาที่อาศัยอยู่ในสภาวะที่มีความเค็มต่างกันยังคงดำเนินต่อไปโดยใช้ตัวอย่างของสารปรุงแต่งรสดั้งเดิมอื่นๆ ได้แก่ กรดซิตริกและซูโครส กรดซิตริกและซูโครสต่างจากเกลือตรงที่ไม่พบในทะเลหรือ น้ำจืดอ่างเก็บน้ำธรรมชาติ พบว่าสำหรับปลาทุกชนิดที่เราใช้ โดยไม่คำนึงถึงความสัมพันธ์กับความเค็ม กรดซิตริกเป็นตัวกระตุ้นรสชาติที่มีประสิทธิภาพสูง สำหรับแมลงสาบ ปลาคาร์พซิลเวอร์ครูเชียน และปลาซิคลาโซมา เซเวอร์รัม รวมถึงปลาลิ้นหมาขั้วโลก กรดซิตริกมีฤทธิ์ขับไล่ คุณสมบัติด้านรสชาติ. ในเวลาเดียวกัน สำหรับปลาดุกลาย ปลาหางนกยูง และปลาก้างปลา 9 หนามนั้น ทำหน้าที่เป็นสารกระตุ้นรสชาติ ส่งผลให้มีการใช้อาหารเม็ดเพิ่มมากขึ้น (ตารางที่ 1) ข้อมูลวรรณกรรมจากปลาสายพันธุ์อื่นๆ ยืนยันผลลัพธ์ของเราว่ากรดซิตริกเป็นสารระคายเคืองต่อรสชาติที่มีประสิทธิภาพสูง ในปลาคาร์พ ถ่านทะเลสาบ ถ่านดาวัตชาน กรดซิตริกทำให้การบริโภคเม็ดเพิ่มขึ้นอย่างรวดเร็ว และในปลาคาร์พขาว ปลาสเตอร์เจียนไซบีเรีย และปลาแซลมอนชุม Opsotropubis keala การบริโภคเม็ดลดลงอย่างรวดเร็ว (Kasumyan, Sidorov, 1992; Kasumyan และคณะ 1993; Kasumyan, Kazhlaev, 1993; KaBituap, 81s1ogou, 1995a, 19956; Kasumyan, Morey, 1996,1997; Kasumyan, 1997) ซูโครสเป็นสารให้รสชาติที่ไม่แยแสสำหรับปลาลิ้นหมาขั้วโลก ปลาดุกลาย Cichlazoma severum ปลาคาร์พ crucian เงิน ปลาสติ๊กหลังเก้าหนาม และทำหน้าที่เป็นสารกระตุ้นรสชาติของแมลงสาบและปลาหางนกยูง การวิเคราะห์ผลลัพธ์ของคุณเองและข้อมูลของผู้อื่น
พันธุ์ปลาที่เราศึกษาแตกต่างกันไม่เพียงแต่ในทัศนคติต่อความเค็มของน้ำเท่านั้น แต่ยังรวมถึงรูปแบบการให้อาหาร องค์ประกอบ และความกว้างของสเปกตรัมอาหารด้วย สิ่งที่น่าสนใจเป็นพิเศษจากมุมมองของการค้นหาความเชื่อมโยงระหว่างนิสัยการกินปลาที่กล่าวข้างต้นกับรสนิยมที่ชอบคือปลาซึ่งมีสัดส่วนอาหารที่มีนัยสำคัญ อาหารจากพืช. จากข้อมูลที่เกี่ยวข้องกับสัตว์มีกระดูกสันหลังกลุ่มอื่นๆ สัตว์ที่กินพืชหรืออาหารผสมมักจะแสดงการตอบสนองต่อรสชาติเชิงบวกต่อโมโน- และไดแซ็กคาไรด์ (Bronstein, 1956; Harborne, 1985) เช่นเดียวกับสารอื่นๆ บางอย่างที่มีรสหวานสำหรับมนุษย์ ( คาสซิล, 1972).
ในบรรดาสายพันธุ์ที่เราใช้ ส่วนประกอบของพืชในอาหารเด่นชัดที่สุดในแมลงสาบ (Domrachev, Pravdin, 1962; Klyuchareva, I960; Grandilevskaya-Dexbach, 1961; Poddubny, 1966; Rodionova, 1969; Wetherley, 1987; Giles et al. , 1990; Horppila, 1994) และ guppies (Dussault และ Kramer, 1981) มันเป็นปลาสองสายพันธุ์จากเจ็ดการศึกษาที่แสดงให้เห็นรสนิยมของซูโครส - ในแมลงสาบการบริโภคเม็ดที่มีสารนี้สูงเป็นสองเท่าของการบริโภคเม็ดควบคุมและในปลาหางนกยูงนั้นสูงกว่า 9 เท่าและ 22 เท่า (สำหรับกลุ่มหมายเลข 1 และหมายเลข 2) อาหารของปลาคาร์พสีเงินยังมีสาหร่ายและแมคโครไฟต์ (Aristovskaya, 1935; Borutsky, 1950; Kharitonova, 1963 เป็นต้น) แต่ซูโครสเป็นสิ่งที่ทำให้ระคายเคืองโดยไม่แยแส ในปลาที่เหลือที่เราศึกษา วัตถุจากพืชในอาหารมีการแสดงออกอย่างอ่อนหรือขาดหายไปโดยสิ้นเชิง และทั้งหมดนี้มีทัศนคติที่ไม่แยแสต่อซูโครส (ตารางที่ 1) เป็นที่ทราบกันดีจากวรรณคดีว่าเดซและปลาคาร์พหญ้าเป็นปลาที่มีอาหารที่มีพืชเป็นส่วนประกอบ (Verigin, 1961; Lupacheva, 1967; Bobrova, 1968;
สตูจ, 1973; โปปอฟ 1975; เวเธอร์ลีย์, 1987) ซูโครสมีคุณสมบัติเป็นสารกระตุ้นรสชาติ ทำให้มีการบริโภคอาหารเม็ดเพิ่มมากขึ้น (Kasumyan และ Moray, 1997; Kasumyan, 1997) สำหรับปลา เช่น ปลาคาร์พ ปลาน้ำจืด ปลาซิว Phoxinus phoxinus ปลาเรนโบว์เทราท์ ปลาชาร์ -
ดาวาชาน, คริสติโวเมอร์ ชาร์, ปลาสเตอร์เจียนไซบีเรีย, ปลาสเตอร์เจียน stellate, ปลาคอด
Gadus morhua, navaga, ปลาเทราท์แคสเปียน และซูโครสปลาแซลมอนชุมเป็นสารปรุงแต่งรสที่ไม่แยแส (Kasumyan, Sidorov, 1992,1993; Kasumyan, Kazhlaev, 1993; Kastimyan, Sidorov, 1995b; Kasumyan, Morey, 1996; Kasumyan, 1997) จากสายพันธุ์ที่ระบุไว้ข้างต้น มีเพียงปลาคาร์พและสร้อยเท่านั้นที่มีพืชอยู่ในอาหาร เช่นเดียวกับปลาคาร์พ crucian เงินที่เราศึกษา แต่ส่วนแบ่งในอาหารของปลาเหล่านี้มีไม่มาก (Stepanova, 1953; Dmitrieva, 1957; Cheremisova, 1958; คาริโตโนวา, 1963; Lebedev, Spanovskaya, 1983) สปีชีส์ที่เหลือไม่ใช้พืชเป็นอาหารหรือกินเป็นระยะๆ โดยมีลักษณะเฉพาะคือการแบ่งสัตว์ในสัตว์
สายพันธุ์ปลาที่เราศึกษายังแตกต่างกันตามความกว้างของรายการอาหารที่พวกเขาบริโภค ดังนั้นตามที่นักวิจัย * หลายคนระบุว่าแมลงสาบเป็นของยูริฟาจ (Zheltenkova, I960; Shmidtov, 1962; Nebolsina, 1965; Grigorash et al., 1973; Ermolin, 1977 เป็นต้น) ปลาคาร์พ crucian เงินยังเป็นของปลากลุ่มเดียวกัน (Aristovskaya, 1935; Dmitrieva, 1957; Stepanova, 1953; Cheremisova, 1958; Kharitonova, 1963) ปลาลิ้นหมาขั้วโลกสามารถจำแนกได้ว่าเป็นสัตว์หน้าดินที่มีสารอาหารหลากหลาย (Voevodin, 1996; Shubnikov et al., 1970; Yu.S. Reshtnikov, การสื่อสารด้วยวาจา), แพลงก์ตอนที่มีอาหารหลากหลาย ได้แก่ ปลาหางนกยูง (Dassault, Kramer, 1981 ) , stickleback เก้าหนาม (Wootton, 1976; Vvedenskaya, 1992, 1993; Maksimenkov, Tokranov, 1994) และ cichlazoma severum (Axelrod, Vorderwinkler, 1993; Pausan, 1984, D.D. Zvorykin, การสื่อสารด้วยวาจา) ปลาดุกลายมีลักษณะเด่นชัดโดย stenophagy (Barsukov, 1983; Karamushko และ Shatunovsky, 1994)
การวิเคราะห์การตอบสนองด้านรสชาติของปลาที่ศึกษาต่อสารปรุงแต่งรสแบบดั้งเดิมแสดงให้เห็นว่าสเปกตรัมทางโภชนาการที่หลากหลายไม่ได้สะท้อนให้เห็นในจำนวนของสารปรุงแต่งรสที่มี คุณสมบัติที่น่าสนใจ. ดังนั้นสารกระตุ้นรสชาติคลาสสิกเพียงชนิดเดียวจาก 4 ชนิดจึงมีผลในการกระตุ้นปลาลิ้นหมาอาร์กติก Cichlazoma severum และปลาตีนติดเก้าหนาม - ปลาที่มีสารอาหารหลากหลายเช่นเดียวกับปลาดุกลาย stenophagous สำหรับยูริฟาจของปลาคาร์พซิลเวอร์ครูเชียน ไม่มีสารที่มีรสชาติดั้งเดิมใดๆ ที่มีผลกระตุ้น ในขณะที่ยูริฟาจของแมลงสาบและปลาหางนกยูงมีสารดังกล่าวสองชนิดในแต่ละครั้ง
ผลลัพธ์ที่คล้ายกันนี้ได้รับเมื่อใช้กรดอะมิโนอิสระเป็นสารแต่งกลิ่นรส ดังนั้น ปลาคาร์พโรชและซิลเวอร์ครูเชียนจึงมีกรดอะมิโนกระตุ้น 8 ตัว ปลาหางนกยูงมี 5 ตัว และปลาลิ้นหมาขั้วโลกมี 0 (ตารางที่ 1) ดังนั้น จำนวนกรดอะมิโนที่น่าสนใจสำหรับปลาสี่ตัวที่เราศึกษาซึ่งมีสเปกตรัมอาหารที่กว้าง แตกต่างกันไปจาก 38 % (ปลาคาร์พแมลงสาบและเงิน crucian) ถึง 0% (ปลาลิ้นหมาอาร์กติก) การที่นักวิจัยคนอื่นมีส่วนร่วมในการวิเคราะห์ผลลัพธ์ยืนยันว่าไม่มีการเชื่อมโยงที่ชัดเจนระหว่างระดับยูริฟากีของปลากับจำนวนกรดอะมิโนที่น่าดึงดูด รสชาติ ดังนั้นในเรนโบว์เทราท์และแซลมอนชุมซึ่งไม่สามารถจัดเป็นยูริฟาจได้จำนวนกรดอะมิโนที่กระตุ้นจะสูงถึง 90% และ 66% ในขณะที่ปลาคาร์พ สร้อย และปลาน้ำจืด ซึ่งมีสารอาหารที่หลากหลายไม่น้อย มีจำนวนเพียง 23%, 19% และ 5% (Kasumyan, 1997) กรดอะมิโนยับยั้ง (9.5%) ถูกค้นพบเฉพาะสำหรับปลาคาร์พ crucian เงิน การมีอยู่ของกรดอะมิโนอิสระไม่เพียงแต่กระตุ้นการบริโภคอาหารเท่านั้น สารที่มีคุณสมบัติยับยั้งรสชาติทำให้สารประกอบประเภทนี้มีบทบาทในการควบคุมโภชนาการของปลาและการกำหนดคุณสมบัติรสชาติได้ชัดเจนยิ่งขึ้น
เมื่อจับเหยื่อได้ ปลาจะตัดสินใจปฏิเสธหรือกลืนอาหาร เป็นที่สนใจที่จะค้นหาว่ามีความสัมพันธ์ระหว่างระดับยูริฟากีในปลากับจำนวนกรดอะมิโนที่มีรสชาติน่ารังเกียจสำหรับปลาหรือไม่ การวิเคราะห์ข้อมูลของเราและข้อมูลวรรณกรรมแสดงให้เห็นว่า ในกรณีของการกระตุ้นกรดอะมิโน ไม่สามารถระบุการเชื่อมต่อดังกล่าวได้ อย่างไรก็ตาม ในความเห็นของเรา แนวโน้มบางอย่างต่อการสำแดงของมันอาจยังคงมีอยู่ ดังนั้น กรดอะมิโนที่มีฤทธิ์ยับยั้งจึงขาดหายไปหรือมีอยู่ในปริมาณเล็กน้อย ไม่เพียงแต่ในปลาที่มีสเปกตรัมอาหารกว้างเท่านั้นที่เราศึกษา (แมลงสาบ - 0%, ปลาหางนกยูง - 0%, ปลาลิ้นหมาขั้วโลก -0%, ปลาคาร์พสีเงิน 9.5% ) แต่ยังอยู่ใน dace (0%), minnow (19%), ปลาสเตอร์เจียนไซบีเรีย (5%), ปลาค็อด (14%) (Kasumyan, Sidorov, 1994a; Kasumyan, 1997) - ปลาที่ใช้สิ่งมีชีวิตเป็นอาหารหลากหลายสำหรับ อาหาร. ในปลาที่มีสเปกตรัมอาหารจำกัด สัดส่วนของกรดอะมิโนยับยั้งอาจสูงกว่า (คริสโตเมียร์ชาร์ - 57%) ในทางกลับกัน กรดอะมิโนจำนวนมากทำให้เกิดการตอบสนองต่อรสชาติที่ยับยั้งในปลาเกรย์ลิงยุโรปชนิดยูริฟาจ (48%)
การวิเคราะห์ความสัมพันธ์ของความชอบด้านรสชาติของสารปรุงแต่งรสแบบคลาสสิกในปลาที่เราศึกษาจากปี 21 ตัวเลือกที่เป็นไปได้การเปรียบเทียบแบบคู่ของ 7 ชนิดเผยให้เห็น 2 กรณีของความสัมพันธ์เชิงลบที่มีนัยสำคัญ และไม่ใช่กรณีเดียวที่มีความสัมพันธ์เชิงบวกที่มีนัยสำคัญ การใช้ข้อมูลการวิเคราะห์จากวรรณกรรมเกี่ยวกับการตอบสนองพฤติกรรมการรับรสต่อสิ่งเร้ารสชาติดั้งเดิมของปลาสายพันธุ์อื่นๆ (รวม 22 ชนิด รวมถึงผลลัพธ์ของเราเอง) แสดงให้เห็นว่าจากการเปรียบเทียบแบบคู่ที่เป็นไปได้ 253 รายการ มีเพียง 25 กรณี (10%) เท่านั้นที่มีความสัมพันธ์ที่มีนัยสำคัญ ถูกระบุ และใน 12 กรณี ความสัมพันธ์นี้เป็นบวก และใน 13 กรณี เป็นลบ จาก 12 คู่สายพันธุ์ที่ระบุความสัมพันธ์เชิงบวก มีเพียง 7 คู่เท่านั้นที่รวมกัน สายพันธุ์น้ำจืด, 2 คู่ประกอบด้วย
ปลาน้ำจืดและปลาทะเล 2 คู่ - จากปลาน้ำจืดและปลาทะเลเพียงคู่เดียวเท่านั้น (ปลาหางนกยูง กลุ่มที่ 1 - navaga) พบความสัมพันธ์เชิงลบที่มีนัยสำคัญสำหรับสายพันธุ์ 13 คู่ ซึ่งประกอบด้วย 6 คู่ ปลาน้ำจืดสามคู่ - จากน้ำจืดและสายพันธุ์ Anadromous สองคู่ - จากน้ำจืดและ สายพันธุ์ทะเล 1 คู่ - จากปลาอพยพและปลาทะเล และ 1 คู่ - จากปลาทะเล การวิเคราะห์ทางสถิติของความชอบด้านรสชาติของกรดอะมิโนอิสระจากการเปรียบเทียบที่เป็นไปได้แบบคู่ของปลา 7 สายพันธุ์ 21 รายการ เผยให้เห็น 2 กรณีมีความสัมพันธ์กันอย่างมีนัยสำคัญ พบความสัมพันธ์เชิงบวกสำหรับปลาหางนกยูงคู่หนึ่ง - ปลาคาร์พ และความสัมพันธ์เชิงลบ - สำหรับปลาหางนกยูงคู่หนึ่ง - ปลาแซลมอนชุม
ผลลัพธ์เหล่านี้แสดงให้เห็นว่าความชอบด้านรสชาติของปลานั้นมีลักษณะเฉพาะโดยมีความเฉพาะเจาะจงของสายพันธุ์ที่แข็งแกร่ง สเปกตรัมการรับรสของสายพันธุ์ที่ศึกษาในกรณีส่วนใหญ่ไม่มีความสัมพันธ์กัน ในกรณีที่มีความสัมพันธ์เชิงบวก คู่มักจะรวมปลาที่มีวิถีชีวิต โภชนาการ หรืออนุกรมวิธานเข้าด้วยกัน ไม่ต้องสงสัยเลยว่าหากต้องการข้อสรุปที่เข้มงวดยิ่งขึ้นเกี่ยวกับธรรมชาติของความสัมพันธ์ที่อยู่ระหว่างการพิจารณา จำเป็นต้องมีการเพิ่มจำนวนพันธุ์ปลาที่ศึกษา แต่ก็สามารถสันนิษฐานได้ว่ามีความเชื่อมโยงโดยตรงระหว่างระดับความเชี่ยวชาญด้านอาหารของปลา หรือ ทัศนคติต่อความเค็มของน้ำและความกว้างของสเปกตรัมในการกระตุ้นรสชาติ เห็นได้ชัดว่าไม่มีสาร 1 รายการ ข้อยกเว้นเพียงอย่างเดียวคือปลาที่มีส่วนประกอบจากพืชที่กำหนดไว้อย่างชัดเจนในอาหารซึ่งมีทัศนคติเชิงบวกต่อซูโครสในขณะที่ซูโครสสำหรับปลาที่เลี้ยงสัตว์ซูโครสมีคุณสมบัติรสชาติที่ไม่แยแส ในความคิดของเรา การขาดความสัมพันธ์ระหว่างความชอบด้านรสชาติและวิถีชีวิตของปลาสามารถอธิบายได้ด้วยความจำเพาะของสายพันธุ์ที่แสดงออกอย่างชัดเจนของความไวต่อรสชาติในปลา ซึ่งแสดงออกแม้ในสายพันธุ์ที่เกี่ยวข้องกันอย่างใกล้ชิดหรือในปลาที่มีระบบนิเวศที่คล้ายคลึงกัน (Kdsumyan, พ.ศ. 2540; ข้อมูลของเรา) ได้รับ
ผลการวิจัยแสดงให้เห็นว่าความจำเพาะของสายพันธุ์ตามความชอบด้านรสชาตินั้นแสดงออกมาในปลาไม่เพียงแต่ในระดับทางเคมีสัมผัสเท่านั้น ดังที่เคยกำหนดไว้ก่อนหน้านี้โดยใช้วิธีอิเล็กโทรสรีรวิทยา (Goob และ Tatiga, 1980; Marsh et al., 1983b; Ishida และ Lshaka, 1987) แต่ ในระดับสิ่งมีชีวิตด้วย เห็นได้ชัดว่าสาเหตุหลักมาจากคุณลักษณะเฉพาะของการรับรสชาติ - ความชอบด้านรสชาติที่มีความจำเพาะสูง - ซึ่งรับประกันความเป็นเอกลักษณ์เฉพาะของสเปกตรัมทางโภชนาการและความแตกต่างของปลาที่อาศัยอยู่ร่วมกัน ประเภทต่างๆตามองค์ประกอบของสิ่งมีชีวิตที่บริโภค (Nikolsky, 1974)
2. อิทธิพลของประสบการณ์อาหารต่อการตอบสนองต่อรสชาติ
วัตถุประสงค์ประการหนึ่งของงานนี้คือเพื่อระบุอิทธิพลที่เปลี่ยนแปลงไปของอาหารที่ปลาบริโภคและประสบการณ์อาหารของแต่ละคนต่อความชอบด้านรสชาติ เราแก้ไขปัญหานี้โดยใช้ตัวอย่างของบุคคลทางจีโนเจเนติกส์ที่เป็นลูกผสมระหว่างปลาทอง (ตัวเมีย) และปลาคาร์พ (ตัวผู้) ซึ่งมีลักษณะเป็นเนื้อเดียวกันทางพันธุกรรมในระดับสูง
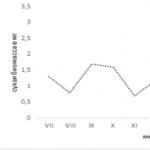
พบว่าลูกผสมที่เติบโตเป็นเวลา 6 เดือนด้วยอาหารที่แตกต่างกัน (อาหารเทียม "Tetramin", tubifex หรือ bloodworm) มีลักษณะเฉพาะที่มีการตอบสนองต่อรสชาติที่คล้ายคลึงกันสูงต่อสารปรุงแต่งรสแบบดั้งเดิมและกรดอะมิโน (รูปที่ 1) เป็นที่ยอมรับกันว่าลูกผสมทุกกลุ่มบริโภคเม็ดที่มีโซเดียมคลอไรด์อย่างเข้มข้นเท่ากันและการบริโภคเม็ดด้วย แคลเซียมคลอไรด์และ กรดมะนาวแตกต่างกันบ้างในหมู่ตัวแทน กลุ่มต่างๆ. ดัชนีนี้มีความคล้ายคลึงกันในระดับสูงระหว่างปลาในกลุ่มต่างๆ ความชอบด้านรสชาติกรดอะมิโน. การวิเคราะห์ความสัมพันธ์ที่ดำเนินการเผยให้เห็นความสัมพันธ์เชิงบวกที่มีนัยสำคัญระหว่างการบริโภคแกรนูลกับสารที่ใช้โดยลูกผสมปลาคาร์พ crucian ของกลุ่มทดลองสามกลุ่ม
การบริโภคเม็ด %
<23456786 группа №1
กลุ่ม Kz2
123456780 กลุ่ม" KsJ
ข้าว. 1. ลิ้มรส ความน่าดึงดูดใจของสารปรุงแต่งรสคลาสสิกและกรดอะมิโนอิสระคัดเลือกสำหรับลูกผสมปลาคาร์พ crucian ที่ปลูกด้วยอาหารเทียม "Tetramin" (กลุ่มที่ 1), Tubifex oligochaetes (กลุ่มที่ 2), "ตัวอ่อน chironomid (กลุ่ม Jfe 3)
1 - โซเดียมคลอไรด์ 1.73M (10%), 2 - แคลเซียมคลอไรด์ 0.9M (10%), 3 - กรดซิตริก 0.26M (5%), 4 - ซูโครส 0.29M (10%), 5 - โพรลีน 0.1M, 6 - ซีรีย์ 0.1M, 7 - อะลานีน 0.1M, 8 - ไกลซีน 0.1M, 9 - การควบคุม
*,*** - ความสำคัญของความแตกต่างที่เกี่ยวข้องกับการควบคุม, น< 0.05 и 0.001."
ความแตกต่างระหว่างกลุ่มปลาที่เปรียบเทียบมีขนาดเล็ก แต่อาจบ่งบอกถึงการเปลี่ยนแปลงในรสชาติของปลาในระหว่างการให้อาหารประเภทต่างๆ ในระยะยาว คุณสมบัติอีกประการหนึ่งคือสารสกัดจากอาหารทั่วไปมีความน่ารับประทานสำหรับปลามากกว่า (ยกเว้นกลุ่มที่ 2 - สารสกัด tubifex มีความน่าดึงดูดน้อยกว่าสารสกัด bloodworm) สิ่งนี้บ่งชี้ว่าการรับประทานอาหารที่เกิดขึ้นไม่เพียงแต่นำไปสู่การเปลี่ยนแปลงในรสนิยมบางอย่างเท่านั้น แต่ยังเพิ่มขึ้นอีกด้วย
พื้นฐานการสะท้อนกลับแบบมีเงื่อนไข ความไวต่อรสชาติต่อสารที่รวมอยู่ในอาหารสัตว์ที่บริโภค
ดังนั้น เมื่อใช้ตัวอย่างของสิ่งเร้ารสชาติคลาสสิกและกรดอะมิโนอิสระ แสดงให้เห็นว่าธรรมชาติของอาหารที่บริโภคมีผลกระทบเล็กน้อยต่อความชอบในรสชาติของปลา สิ่งนี้บ่งชี้ว่าการรับรู้ทางเคมีของรสชาติในปลามีความเป็นพลาสติกที่ค่อนข้างต่ำ โดยสัมพันธ์กับปัจจัยทางโภชนาการที่สำคัญ เช่น องค์ประกอบของอาหาร และยังแสดงให้เห็นถึงการกำหนดรสนิยมทางพันธุกรรมในระดับสูงอีกด้วย ผลลัพธ์ของเราเกี่ยวกับอิทธิพลเล็กน้อยของประสบการณ์ด้านอาหารต่อความชอบด้านรสชาติของปลาได้รับการยืนยันจากข้อมูลทางวรรณกรรม (Kasumyan, Morsy, 1997; Kasumyan, Morsy, 1998) ตามที่ปลาคาร์พหญ้าวัยอ่อนเลี้ยงเป็นเวลา 6 เดือนกับสัตว์ (bloodworm) หรือ พืช (แหน Lemna minor, ผักกาดหอม Lactuca sativa) เข้มงวดมีลักษณะคล้ายคลึงกันอย่างมีนัยสำคัญในการตั้งค่ารสชาติ สำหรับสัตว์มีกระดูกสันหลังกลุ่มอื่นๆ มีการยอมรับว่าความชอบด้านรสชาติอยู่ภายใต้การควบคุมทางพันธุกรรมอย่างเข้มงวด (Ramirez และ Sprott, 1978; Lush, 1981; Shingai และ Beidler, 1985; Sinclair et al., 1992)
การเปรียบเทียบสเปกตรัมรสชาติของลูกผสมปลาคาร์พ crucian และสายพันธุ์พ่อแม่ทำให้เราสามารถประเมินธรรมชาติของการสืบทอดความชอบในรสชาติของปลาได้ ผลการวิจัยพบว่าในแง่ของการตอบสนองด้านรสชาติ ปลาลูกผสมนั้นใกล้กับปลาคาร์พมากกว่าปลาคาร์พ crucian สีเงินมาก (ข้อมูลเกี่ยวกับปลาคาร์พนำมาจาก Kasumyan และ Morey, 1996) ความคล้ายคลึงนี้ชัดเจนเป็นพิเศษเมื่อเปรียบเทียบการตอบสนองด้านรสชาติของปลากับซิตริก กรด แคลเซียมคลอไรด์ และสารอื่นๆ บางชนิด ดังนั้น สารปรุงแต่งรสแบบดั้งเดิมระดับการบริโภคของเม็ดที่มีกรดซิตริกจึงต่ำที่สุดในปลาคาร์พ crucian เงิน ในขณะที่ในลูกผสมและปลาคาร์พ กรดซิตริกเป็นหนึ่งในสารที่น่าสนใจที่สุดใน รสชาติ.
การตอบสนองด้านรสชาติระหว่างสายพันธุ์ดั้งเดิมและลูกผสมทำให้เราสามารถพูดเกี่ยวกับการสืบทอดประเภท patroclinic ของความชอบด้านรสชาติของสารระหว่างการผสมข้ามพันธุ์ ข้อสรุปนี้ได้รับการยืนยันโดยการคำนวณทางสถิติ ซึ่งทำให้สามารถตรวจจับความสัมพันธ์เชิงบวกของการตอบสนองต่อรสชาติต่อกรดอะมิโนและสารแต่งกลิ่นแบบดั้งเดิมระหว่างปลาคาร์พและลูกผสม (r5 = 0.51, p< 0.01). Достоверной связи между ответами серебряного карася и гибридов не установлено (rs = - 0.08, р > 0.05).
3. ศึกษาพฟิสซึ่มทางเพศของรสนิยมในปลา
หนึ่งในงานวิจัยที่ยังไม่ได้รับการพัฒนาก่อนหน้านี้เกี่ยวกับการรับรู้รสชาติคือคำถามเกี่ยวกับการมีอยู่ของความหลากหลายทางเพศในรสนิยมในรสชาติของปลา จำนวนการศึกษาที่มีหัวข้อการศึกษาเพื่ออธิบายความแตกต่างทางเพศในโภชนาการปลานั้นมีน้อยมาก (Baird, 1965; Dussault and Kramer, 1981; Sakashita, 1992; Aburto-Oropeza et al., 2000; Laland and Reader, 1999) จากข้อมูลเหล่านี้ โภชนาการของปลาตัวเมียและตัวผู้ที่อยู่ใน biotopes เดียวกันอาจแตกต่างกัน ซึ่งบ่งชี้ถึงความคลาดเคลื่อนในความชอบด้านรสชาติของปลาที่มีเพศต่างกัน
ปลาหางนกยูงชนิดหนึ่งที่พบความแตกต่างทางเพศในด้านโภชนาการคือปลาหางนกยูง (Dussault and Kramer, 1981) งานวิจัยนี้มีวัตถุประสงค์เพื่อศึกษาเปรียบเทียบความชอบและคุณลักษณะของการแสดงการตอบสนองทางพฤติกรรมต่อสารปรุงแต่งรสชนิดต่างๆ ในเพศตรงข้าม โดยใช้ปลาสายพันธุ์นี้เป็นตัวอย่าง ในการทดลองใช้สารปรุงแต่งรสหลายชนิด ได้แก่ สารกระตุ้น (ซูโครส กรดซิตริก ไกลซีน กรดแอล-กลูตามิก) สารยับยั้ง (แอล-ฮิสทิดีน) และ
ปริมาณการใช้เม็ด %
1 00 90 8 070 -6 05 0403 020 -1 0 -
■ชาย 1; ส เอม เค ฉัน
1-G-T-1-G»T-I-g-t-g-
สิ่งกระตุ้นทางเคมี
ข้าว. 2. จัดอันดับชุดสิ่งเร้าทางเคมี รวบรวมตามขนาดของความน่าดึงดูดใจของปลาหางนกยูงตัวผู้และตัวเมีย
1 - การควบคุม, 2 - แคลเซียมคลอไรด์ 0.9M (10%), 3 - โซเดียมคลอไรด์ 1.73M (10%), A - กรดซิตริก 0.26M (5%), 5 - b-histidine 0.1M, 6 - b-glutamic กรดกรด 0.01M, 7 - ซูโครส 0.29M (10%), 8 - ไกลซีน 0.1M, 9 - สารสกัดไคโรโนมิด 75 กรัม/ลิตร
**. *** - ความสำคัญของความแตกต่าง น< 0.05, 0.01, 0.001 соответственно, до косой черты - по отношению к контролю, после косой черты - между самцами и самками.
ไม่แยแส (โซเดียมคลอไรด์, แคลเซียมคลอไรด์) ผลลัพธ์ของเราแสดงให้เห็นว่าปลาหางนกยูงไม่มีความแตกต่างทางเพศที่เด่นชัดในด้านรสนิยม สำหรับปลาหางนกยูงตัวผู้และตัวเมีย สารชนิดเดียวกันที่ใช้มีประสิทธิผลด้านรสชาติและ
ลำดับของสารตามลำดับที่สร้างขึ้น
แยกกันสำหรับบุคคลทั้งสองเพศ เกือบจะเหมือนกันทั้งหมด (รูปที่ 2) ความคล้ายคลึงกันที่ชัดเจนของสเปกตรัมรสชาติยังระบุด้วยค่าสัมประสิทธิ์สหสัมพันธ์ระหว่างค่าที่สูง
การบริโภคปลาหางนกยูงตัวผู้และตัวเมียที่มีสารที่มีความอร่อยต่างกันไป (r5 = 0.97; p< 0.01). Поскольку у большинства видов рыб, в отличие от гуппи, отсутствует ярко выраженный половой диморфизм в морфологии, размерах и поведении, в частности, пищевом, вполне справедливым представляется предположение о том, что для этих рыб в целом, по-видимому, не характерен половой диморфизм по их вкусовым предпочтениям.
ความแตกต่างระหว่างปลาหางนกยูงตัวเมียและตัวผู้เกี่ยวข้องเฉพาะความรุนแรงของการสำแดงองค์ประกอบแต่ละอย่างของการตอบสนองต่อรสนิยมทางพฤติกรรมเท่านั้น ดังนั้น จึงเผยให้เห็นความแตกต่างที่มีนัยสำคัญระหว่างเพศชายและเพศหญิงในลักษณะต่างๆ เช่น จำนวนการจับและระยะเวลาในการจับเม็ดหลังจากการจับครั้งแรกและตลอดการทดลอง ความแตกต่างเหล่านี้ส่งผลกระทบเฉพาะความรุนแรงของการสำแดงองค์ประกอบเหล่านี้ของการตอบสนองต่อรสชาติเชิงพฤติกรรม ซึ่งในปลา เช่น ปลาหางนกยูง ซึ่งเป็นปลาที่มีชีวิตชีวาและมีลักษณะเฉพาะโดยพฟิสซึ่มทางเพศที่เด่นชัดในขนาดลำตัว อาจเกี่ยวข้องกับลักษณะเฉพาะของชีววิทยาของสายพันธุ์นี้ (Dussault, Kramer, 1981; Laland, Reader, 1999)
4. ลักษณะเฉพาะของการตอบสนองพฤติกรรมต่ออาหารเม็ดในปลาที่มีไลฟ์สไตล์ต่างกัน
การตอบสนองเชิงพฤติกรรมของปลาต่อเม็ดที่นำเสนอนั้นค่อนข้างเรียบง่ายและมีองค์ประกอบพื้นฐานหลายประการ ได้แก่ การจับและการปฏิเสธเม็ด การจับซ้ำ ๆ การจับเม็ดในช่องปาก และในบางกรณี การเคี้ยวเม็ด เช่นเดียวกับในปลาเทราท์สีน้ำตาล (คาสุมยาน) และ Sidorov, 1993, 1994b) ความถี่หรือความเข้มข้นของการแสดงออกขององค์ประกอบเหล่านี้โดยปลาจะแตกต่างกันไปและขึ้นอยู่กับวิถีชีวิตของปลาและคุณสมบัติด้านรสชาติของวัตถุที่เป็นอาหาร (Kasumyan, 1997)
สารปรุงแต่งรสแบบคลาสสิกและกรดอะมิโนอิสระของปลาทั้งหมดที่เราศึกษาในปลาแมลงสาบ ปลาคาร์พสีเงิน และปลา Cichlazoma severum เผยให้เห็นความสัมพันธ์เชิงลบที่มีนัยสำคัญระหว่างจำนวนเหตุการณ์การจับและความน่าดึงดูดของปลาเม็ด ยิ่งความน่าดึงดูดใจของเม็ดปลาสำหรับปลาเหล่านี้สูงเท่าใด การจับปลาก็จะน้อยลงตามความสัมพันธ์กับเม็ดเล็กด้วยการกระตุ้นนี้ นี่เป็นการยืนยันข้อสรุปก่อนหน้านี้ว่าเม็ดปลากลืนซึ่งน่าดึงดูดใจในการลิ้มรสเป็นส่วนใหญ่ในครั้งแรก (Kasumyan, 1997) อย่างไรก็ตาม สำหรับปลาหางนกยูงจะสังเกตเห็นภาพที่ตรงกันข้าม - ยิ่งความน่าดึงดูดใจของสิ่งเร้ายิ่งสูงเท่าใดจำนวนการกระทำที่จับก็จะยิ่งมากขึ้นเท่านั้น ดำเนินการโดยปลาทดลอง ข้อมูลที่มีอยู่เกี่ยวกับสายพันธุ์อื่นแสดงให้เห็นว่าปลาแม่น้ำหลายชนิด (สร้อย ปลาน้ำจืด ปลาน้ำจืด ปลาเกรย์ยุโรป ปลาเทราท์สีน้ำตาล ปลาเรนโบว์เทราต์ ฯลฯ) ในระหว่างการทดลองค่อนข้างจะไม่ค่อยปฏิเสธแล้วจับปลาเม็ดใหม่ (Kasumyan, Sidorov, 1993, 19946; Kavituap, B Sigory, 1995a; Kasumyan, 1997) ลักษณะพิเศษของการตอบสนองต่อรสชาติเชิงพฤติกรรมของปลาเหล่านี้มีความเกี่ยวข้องกับถิ่นที่อยู่ของพวกมันในบริเวณแม่น้ำและลำธารที่มีอัตราการไหลสูงอย่างไม่ต้องสงสัย ภายใต้เงื่อนไขดังกล่าว รายการอาหารที่ถูกปฏิเสธจะถูกลำเลียงไปตามกระแสน้ำอย่างรวดเร็วและไม่สามารถเข้าถึงได้ โดยเฉพาะอย่างยิ่งสำหรับปลาที่มีวิถีชีวิตในอาณาเขตและไม่ได้ออกล่าเหยื่อเป็นเวลานาน ในปลา (ปลาคาร์พ ฯลฯ) ที่อาศัยอยู่เป็นส่วนใหญ่ในน้ำไหลช้าหรือน้ำนิ่ง เช่นเดียวกับในปลาทะเล เช่น ปลาคอดและปลาคอดสีเหลือง การจับเม็ดปลาซ้ำๆ จะเกิดขึ้นบ่อยกว่ามาก (Kasumyan และ Morey, 1996;
กัสสุมยาน, 2540) อย่างไรก็ตาม ไม่เหมือนกับปลาค็อดและนาวากา
ปลาลิ้นหมาขั้วโลกที่เราศึกษาแสดงให้เห็นความสัมพันธ์แบบผกผัน ซึ่งอาจเนื่องมาจากถิ่นที่อยู่ของมันในบริเวณชายฝั่งทะเลทางตอนเหนือ ซึ่งมีกระแสน้ำไหลแรงเป็นพิเศษ
ในการตอบสนองรสชาติของแมลงสาบ ปลาทอง ปลาหางนกยูง ปลาลิ้นหมาอาร์กติก และปลาลิ้นหมาหนามเก้าหนามต่อสิ่งเร้ารสชาติคลาสสิกและกรดอะมิโน พบว่ามีความสัมพันธ์เชิงบวกระหว่างความอร่อยของอาหารเม็ดและระยะเวลาในการคงอยู่ในช่องปาก ปลาเหล่านี้มีลักษณะเฉพาะคือการทดสอบเม็ดด้วยสารที่น่าดึงดูดต่อรสชาตินานขึ้น ก่อนหน้านี้ (Kasumyan และ Morey, 1996) มีข้อเสนอแนะว่าคุณลักษณะของการตอบสนองต่อรสชาตินี้อาจเกี่ยวข้องกับการให้อาหารปลาหน้าดินซึ่งต้องใช้เวลานานในการแยกอาหารออกจากเศษซากจำนวนมากที่จับมาพร้อมกับอาหาร8 "1 พันล้านปอนด์, 1991) ผลลัพธ์ของเราแสดงให้เห็นว่าความสัมพันธ์ที่คล้ายคลึงกันอาจมีลักษณะเฉพาะไม่เพียงกับสัตว์หน้าดินเท่านั้น แต่ยังรวมถึงแพลงก์ติวอร์ด้วย เช่นเดียวกับในกรณีของ Sticklebacks และ Guppies ที่มีหนามเก้าหนาม การระบุสารที่ดึงดูดต่อรสชาติได้อย่างชัดเจน มั่นใจ และไม่มีข้อผิดพลาด พร้อมด้วยการทดสอบรสชาติซ้ำๆ ของเม็ดในบางกรณี และการเริ่มต้นการสะท้อนกลับของการกลืนอาหารโดยใช้ข้อมูลดังกล่าวต้องใช้เวลานานกว่า . การปฏิเสธอาหารที่รวดเร็วขึ้นโดยมีคุณสมบัติด้านรสชาติที่ไม่พึงปรารถนาสามารถอธิบายได้โดยการลดเวลาที่ไม่ก่อให้เกิดผลที่ใช้ไประหว่างการให้อาหารให้เหลือน้อยที่สุด อย่างไรก็ตาม ข้อสันนิษฐานดังกล่าวไม่เป็นสากล เนื่องจาก ตัวอย่างเช่น ในปลาแซลมอนชุม ความสัมพันธ์ระหว่างระยะเวลาในการทดสอบรสชาติอาหารและความน่าดึงดูดของมัน ซึ่งแตกต่างจากสายพันธุ์อื่น ๆ เป็นสิ่งที่ตรงกันข้าม (Kasumyan, Sidorov, 1992) ผู้เขียนหลายคนแนะนำว่า > การเก็บรักษาวัตถุอาหารในช่องปากในระยะยาวสามารถช่วยให้ประเมินรสชาติได้แม่นยำยิ่งขึ้น (Tamar, 1976; Kasumyan, Sidorov, 1993, 19946)
พบว่าการตอบสนองด้านรสชาติของปลาคาร์พพันธุ์ผสม crucian ที่ปลูกด้วยอาหารที่แตกต่างกันมีลักษณะเฉพาะสูง
ความคล้ายคลึงกันไม่เพียง แต่ในระดับการบริโภคของเม็ดชนิดเดียวกันเท่านั้น แต่ยังรวมถึงความบังเอิญของรูปแบบทั่วไปของการตอบสนองต่อรสชาติ - การปฏิเสธจำนวนเล็กน้อยและการจับเม็ดซ้ำ ๆ ซ้ำ ๆ ซึ่งเป็นความสัมพันธ์เชิงบวกที่เด่นชัดระหว่างความน่าดึงดูดใจของรสชาติ เม็ดและระยะเวลาการเก็บรักษาในช่องปาก สิ่งนี้บ่งชี้ว่าประสบการณ์การให้อาหารที่แตกต่างกันไม่ส่งผลต่อการแสดงออกขององค์ประกอบพื้นฐานของการตอบสนองต่อรสชาติของปลา
บทสรุป
ผลการศึกษาพบว่าปลาที่มีระบบนิเวศต่างกันมีการรับรู้รสชาติที่ได้รับการพัฒนามาอย่างดี ทำให้สามารถแยกแยะคุณสมบัติรสชาติของสารประเภทต่างๆ ได้ การตอบสนองด้านรสชาติของปลานั้นถูกกำหนดโดยพันธุกรรมอย่างเคร่งครัด โดยในระหว่างการผสมข้ามพันธุ์ระหว่างปลาคาร์พ Silver Crucian และปลาคาร์พ พวกมันจะได้รับการสืบทอดตามประเภท Patroclinic ความเชื่อมโยงระหว่างความชอบด้านรสชาติของปลากับสภาพที่อยู่อาศัยหรือ
ตรวจไม่พบลักษณะของสารอาหารเนื่องจากเห็นได้ชัดว่ามีความจำเพาะของสายพันธุ์ในระดับสูงของสเปกตรัมรสชาติซึ่งแสดงออกไม่เพียง แต่ในทางเคมีเท่านั้น แต่ยังรวมถึงระดับสิ่งมีชีวิตด้วย คุณลักษณะนี้แสดงให้เห็นอย่างชัดเจนถึงความสำคัญและแน่นอนในการกำหนดบทบาทของการรับรู้รสชาติในการรับประกันการเลือกสรรโภชนาการของปลา รวมถึงความสามารถในการเลือกบริโภครายการอาหารที่เป็นลักษณะเฉพาะของพวกเขา
ข้อมูลที่ได้รับพร้อมกับข้อมูลที่มีอยู่เกี่ยวกับการไม่มีความจำเพาะของประชากรที่เห็นได้ชัดเจนในเรื่องความชอบด้านรสชาติของปลา (Kasumyan, Sidorov, 1995) และการไม่มีอิทธิพลที่เห็นได้ชัดเจนของประสบการณ์การให้อาหารปลาแต่ละตัว (Kasumyan, Morey, 1997; KaBituap , Mogeu, 1998; ข้อมูลของเรา) ให้เหตุผลในการยืนยันว่าสเปกตรัมรสชาติในปลาถือได้ว่าเป็นลักษณะเฉพาะของประชากรเหนือสายพันธุ์เฉพาะ โดยไม่ขึ้นอยู่กับเพศหรืออาหารที่บริโภค
รูปแบบการรับรู้รสชาติของปลาหลายรูปแบบและลักษณะเฉพาะของการตอบสนองต่อสารปรุงแต่งชนิดต่างๆ ถือเป็นประโยชน์ในทางปฏิบัติที่สำคัญ และสามารถนำมาใช้ในการแก้ปัญหาต่างๆ ในการประมงและการเพาะเลี้ยงสัตว์น้ำ การศึกษาที่ดำเนินการแสดงให้เห็นถึงคำมั่นสัญญาในการค้นหาและสร้างสารกระตุ้นและสารยับยั้งที่มีประสิทธิภาพสูงสำหรับปลา และทำหน้าที่เป็นพื้นฐานทางชีวภาพสำหรับการพัฒนาวิธีการควบคุมพฤติกรรมการให้อาหารปลาโดยใช้สิ่งกระตุ้นรสชาติ ผลลัพธ์ที่เราได้รับสามารถนำมาใช้เพื่อเพิ่มความอร่อยของอาหารสัตว์ ดำเนินการแก้ไขสูตรโดยการเติมสารพิเศษที่มีฤทธิ์กระตุ้นสูง หรือแยกส่วนประกอบที่มีสารประกอบยับยั้งออกจากอาหารสัตว์ ซึ่งไม่เพียงแต่จะลดน้อยลงเท่านั้น
การสูญเสียอาหารเทียมโดยตรงในระหว่างการเพาะปลูก แต่ยังช่วยให้การแปลงอาหารเพื่อการเจริญเติบโตของปลามีประสิทธิภาพมากขึ้นด้วย เนื่องจากเป็นที่ทราบกันดี (Takea และ Takp, 1992) ว่าการบริโภคอาหารที่น่าสนใจด้วยเคมีเซ็นเซอร์จะมาพร้อมกับปลาโดยการหลั่งของระบบย่อยอาหารที่รุนแรงมากขึ้น เอนไซม์
1. ปลาที่ทำการศึกษาในระบบนิเวศที่แตกต่างกันมีการตอบสนองพฤติกรรมด้านรสชาติที่ชัดเจนต่อสารเคมีประเภทต่างๆ - กรดอะมิโนอิสระและสารรับรสดั้งเดิม ได้รับการยืนยันว่ามีความจำเพาะของสายพันธุ์ปลาในระดับสูง
2. ไม่มีความสัมพันธ์ระหว่างความชอบในรสชาติของปลากับทัศนคติต่อความเค็มของน้ำ ปลาทะเล ปลาน้ำจืด และปลายูริฮาลีนไม่มีความกว้างและองค์ประกอบของสเปกตรัมของสารที่แตกต่างกัน โดยมีคุณสมบัติในการกระตุ้น ไม่แยแส หรือยับยั้งรสชาติ
3. ไม่พบความสัมพันธ์ระหว่างลักษณะการให้อาหารหลักของปลา (ธรรมชาติของสารอาหาร ความกว้างของช่วงของสิ่งมีชีวิตในอาหารที่บริโภค) และรสนิยมของปลา (จำนวนสารปรุงแต่งรสที่กระตุ้น ส่วนประกอบ)
4. เป็นที่ยอมรับแล้วว่าปลาที่มีส่วนประกอบจากพืชที่กำหนดไว้อย่างชัดเจนในอาหารนั้นมีทัศนคติเชิงบวกต่อซูโครส สำหรับปลาที่เลี้ยงสัตว์ซูโครสมีคุณสมบัติด้านรสชาติที่ไม่แยแส
5. ความชอบด้านรสชาติของปลานั้นถูกกำหนดโดยพันธุกรรมอย่างเคร่งครัด ประสบการณ์การรับประทานอาหารของแต่ละบุคคลมีผลในการปรับเปลี่ยนรสชาติของปลาเล็กน้อย การสืบทอดความชอบด้านรสชาติของปลา
การผสมข้ามพันธุ์แบบเฉพาะเจาะจง (ปลาคาร์พ crucian สีทอง, ตัวเมีย x ปลาคาร์พ, ตัวผู้) มีลักษณะเป็น patroclinic
6. รสนิยมความชอบของปลาหางนกยูงไม่มีความแตกต่างทางเพศ ความแตกต่างระหว่างบุคคลที่มีเพศต่างกันเกี่ยวข้องเฉพาะความรุนแรงของการแสดงออกขององค์ประกอบแต่ละส่วนของการตอบสนองพฤติกรรมการรับรสซึ่งสะท้อนถึงลักษณะเฉพาะของชีววิทยาของการสืบพันธุ์ของปลาหางนกยูง
7. แสดงให้เห็นว่าลักษณะเชิงปริมาณขององค์ประกอบหลักในการตอบสนองรสชาติเชิงพฤติกรรม (จำนวนการจับของเม็ด ระยะเวลาในการคงของเม็ดในช่องปาก) แตกต่างกันในปลาที่มีระบบนิเวศต่างกัน ในปลาสายพันธุ์ส่วนใหญ่ที่ศึกษา มีความสัมพันธ์เชิงลบระหว่างจำนวนการจับวัตถุอาหารซ้ำๆ และความอร่อยของมัน จำนวนการจับที่น้อยที่สุดนั้นพบได้ในปลารีโอฟิลิก ปลาหน้าดินส่วนใหญ่มีลักษณะการกักเก็บวัตถุอาหารในช่องปากได้นานขึ้น ไม่พบการเปลี่ยนแปลงประสบการณ์ด้านอาหารต่อการตอบสนองพฤติกรรมด้านรสชาติของปลา
1. Kasumyan A.O., Sidorov S.S., Platova-Nikolaeva E.V. พ.ศ. 2536 การตอบสนองเชิงพฤติกรรมของปลาหมาป่ารุ่นเยาว์ต่อสิ่งเร้ารสชาติคลาสสิก // ใน: บทคัดย่อของเอกสารทางวิทยาศาสตร์และโปสเตอร์สำหรับการประชุมสัมมนาเรื่องการเลี้ยงปลารุ่นเยาว์จำนวนมาก เบอร์เกน นอร์เวย์ 21-23 มิถุนายน 2536 โคเปนเฮเกน ป.37.
2. Kasumyan A.O., Platova-Nikolaeva E.B. พ.ศ. 2538 ศึกษาความไวต่อรสชาติของปลาบางชนิดในทะเลสีขาวที่มีทัศนคติต่อความเค็มของน้ำที่แตกต่างกัน // ใน: ปัญหา
การศึกษา การใช้เหตุผล และการคุ้มครองทรัพยากรธรรมชาติของทะเลสีขาว เซนต์ปีเตอร์สเบิร์ก. ซิน ราส. หน้า 122-123.
3. Kasumyan.A.O., Marusov E.A., Nikolaeva E.V..1996 อิทธิพลของประสบการณ์ทางอาหารต่อรสชาติและความไวในการรับกลิ่นของปลา // ในคอลเลกชัน: พฤติกรรมของปลา บทคัดย่อรายงาน อันดับที่ 2 รัสเซียทั้งหมด ประชุม โบร็อก หน้า 35-36.
4. Kasumyan A.O., Nikolaeva E.V. พ.ศ. 2540 ความชอบด้านรสชาติของปลาหางนกยูง Poecilia reticulata (Cyprinodontiformes, Pisces) // ปัญหาเกี่ยวกับวิทยาวิทยา ต. 37 ลำดับ 5 หน้า 696-703
5. Kasumyan A.O., Marusov E.A., Morsy A.M.N., Nikolaeva E.V. พ.ศ. 2541 การเปรียบเทียบการตอบสนองทางพฤติกรรมการรับกลิ่นและรสชาติต่อกรดอะมิโนอิสระในปลาไซปรินิดสองตัว // ความรู้สึกทางเคมี V. 23, N 5, P. 642-643. "
6. Nikolaeva E.B., Kasumyan A.O. 2543. การวิเคราะห์เปรียบเทียบความชอบด้านรสชาติและการตอบสนองต่อพฤติกรรมต่อสิ่งเร้ารสชาติในปลาหางนกยูงตัวเมียและตัวผู้ Poecilia reticulata // คำถามเกี่ยวกับ Ichthyology ต.40 ลำดับที่ 4.ป.560-565.
บทที่ 1 การทบทวนวรรณกรรม
บทที่ 2 วัสดุและวิธีการ
2.1. วัตถุประสงค์ของการวิจัย
2.2. เงื่อนไขการกักขังและการเตรียมปลาทดลองเพื่อการทดลอง
2.3. ระเบียบวิธีในการทำการทดลองเชิงพฤติกรรม
2.4. วิธีเตรียมเม็ดวุ้น-วุ้นทดลอง
2.5. ลักษณะทั่วไปของวัสดุ
บทที่ 3 ผลลัพธ์
3.1. ศึกษาการตอบสนองพฤติกรรมการรับรสของปลาจากระบบนิเวศต่างๆ ต่อสารปรุงแต่งรสดั้งเดิมและกรดอะมิโนอิสระ
3.1.1. แมลงสาบ
3.1.2. ปลาทอง.
3.1.3. ปลาหางนกยูง
3.1.4. Cichlazoma severum
3.1.5. Stickleback เก้าหนาม
3.1.6. ปลาลิ้นหมาอาร์กติก
3.1.7. ปลาดุกลาย.
3.2. ลิ้มรสการตอบสนองทางพฤติกรรมของปลาคาร์พพันธุ์ผสม crucian ที่ปลูกโดยใช้อาหารที่แตกต่างกัน
3.3. การตอบสนองพฤติกรรมการกินของปลาหางนกยูงตัวผู้และตัวเมีย
บทที่ 4 การอภิปรายผล 54 บทสรุป 90 รายชื่อวรรณกรรมที่อ้างถึง. 92 ภาคผนวก
การแนะนำ วิทยานิพนธ์ทางชีววิทยา ในหัวข้อ “การศึกษาความชอบด้านรสชาติของปลา”
การรับรู้รสชาติมีบทบาทสำคัญในการสนับสนุนทางประสาทสัมผัสในช่วงสุดท้ายของพฤติกรรมการให้อาหารที่เกี่ยวข้องกับการประเมินคุณสมบัติรสชาติของเหยื่อและความสอดคล้องกับความต้องการทางโภชนาการของปลา และการใช้รายการอาหารที่เพียงพอ การศึกษาระบบการรับรสของปลาจำนวนมากที่ดำเนินการจนถึงปัจจุบัน มุ่งเน้นไปที่การอธิบายลักษณะทางสัณฐานวิทยาและภูมิประเทศของปุ่มรับรส โครงสร้างพิเศษและการปกคลุมด้วยเส้น และการศึกษาโครงสร้างทางสัณฐานวิทยาของส่วนกลาง การศึกษาคุณสมบัติการทำงานของระบบรสชาติในกรณีส่วนใหญ่จะดำเนินการโดยใช้วิธีอิเล็กโทรสรีรวิทยา วิธีการประเมินความชอบด้านรสชาติของปลายังคงพัฒนาได้ไม่ดีและเป็นผลให้ข้อมูลเกี่ยวกับทัศนคติของปลาต่อรสชาติของสารประเภทต่างๆ แทบไม่มีเลยจนกระทั่งเมื่อไม่นานมานี้ ปริมาณข้อมูลการทดลองที่ได้รับโดยใช้ปฏิกิริยาทดสอบพฤติกรรมและเผยให้เห็นถึงรสชาติที่ชอบของปลานั้นมีจำกัดและไม่ได้ให้แนวคิดที่ชัดเจนเกี่ยวกับรูปแบบทั่วไปและคุณลักษณะเฉพาะของทัศนคติของปลาต่อรสชาติสิ่งเร้า ความเหมือนและความแตกต่างใน สเปกตรัมของสารปรุงแต่งรสที่มีประสิทธิผลในปลาที่มีวิถีชีวิตและตำแหน่งที่แตกต่างกัน
อิทธิพลต่อความไวต่อรสชาติของปลาของปัจจัยทางชีวภาพและปัจจัยทางชีวภาพต่างๆ สภาพความเป็นอยู่ของปลา และลักษณะของโภชนาการของปลายังคงมีการศึกษาที่ไม่ดีนัก คำถามเกี่ยวกับการมีอยู่และธรรมชาติของการเชื่อมโยงระหว่างความชอบด้านรสชาติกับความกว้างของสเปกตรัมอาหารของปลา องค์ประกอบของสิ่งมีชีวิตในอาหารที่บริโภค และประสบการณ์ทางโภชนาการของแต่ละบุคคลยังคงเปิดกว้างอย่างสมบูรณ์ ไม่มีข้อมูลเกี่ยวกับความรุนแรงของพฟิสซึ่มทางเพศในรสนิยมในปลาหรือเกี่ยวกับอิทธิพลของวิถีชีวิตของปลาที่มีต่อการสำแดงองค์ประกอบต่าง ๆ ของการตอบสนองเชิงพฤติกรรมต่อสิ่งเร้ารสชาติ
การศึกษาประเด็นเหล่านี้และประเด็นเฉพาะอื่นๆ ของการรับรู้รสชาติในปลามีความสำคัญทางทฤษฎีที่สำคัญ เนื่องจากทำให้สามารถชี้แจงบทบาทของระบบประสาทสัมผัสนี้ในการเลือกวัตถุอาหารที่เพียงพอโดยปลาได้ ความรู้เกี่ยวกับรูปแบบของความไวต่อรสชาติของปลาคุณสมบัติเฉพาะของการตอบสนองต่อสารปรุงแต่งประเภทต่าง ๆ นั้นเป็นที่สนใจในทางปฏิบัติอย่างไม่ต้องสงสัยและสามารถนำมาใช้ในการแก้ปัญหาต่าง ๆ ในการประมงและการเพาะเลี้ยงสัตว์น้ำ
งานวิจัยนี้มีวัตถุประสงค์เพื่อศึกษาลักษณะความชอบด้านรสชาติของปลา วัตถุประสงค์ของงานวิจัย ได้แก่ การเปรียบเทียบความชอบด้านรสชาติของปลาที่อาศัยอยู่ในน้ำที่มีความเค็มต่างกัน ศึกษาความสัมพันธ์ระหว่างรูปแบบการให้อาหาร สเปกตรัมอาหาร และความชอบด้านรสชาติของปลา ศึกษาอิทธิพลของประสบการณ์ด้านอาหารต่อความชอบด้านรสชาติของปลา การกำหนดลักษณะการสืบทอดความชอบด้านรสชาติของปลา การศึกษาความหลากหลายทางเพศของรสนิยมในปลา (โดยใช้ตัวอย่างของปลาหางนกยูง) การอธิบายลักษณะของการสำแดงองค์ประกอบหลักของการตอบสนองพฤติกรรมรสชาติของปลาในระบบนิเวศที่แตกต่างกัน
ความแปลกใหม่ทางวิทยาศาสตร์ของวิทยานิพนธ์นี้อยู่ที่การขยายความเข้าใจเกี่ยวกับลักษณะการทำงานของระบบรสชาติของปลา ผลงานเผยเป็นครั้งแรกถึงรสชาติของปลา 7 สายพันธุ์สำหรับสารปรุงแต่งรสคลาสสิก และปลา 4 สายพันธุ์สำหรับกรดอะมิโนอิสระ เป็นครั้งแรกที่มีการกำหนดจุดยืนเกี่ยวกับการไม่มีความสัมพันธ์ระหว่างความชอบด้านรสชาติของปลากับทัศนคติต่อความเค็มของน้ำ ระหว่างระดับยูริฟากีของปลาและความกว้างของสเปกตรัมของสารกระตุ้น พบว่าปลาที่มีส่วนประกอบจากพืชชัดเจนมีทัศนคติเชิงบวกต่อน้ำตาล (ซูโครส)
นับเป็นครั้งแรกที่การใช้บุคคลที่มีลักษณะคล้ายไอโซเจนิกที่เลี้ยงด้วยอาหารประเภทต่างๆ แสดงให้เห็นว่าความชอบด้านรสชาติของปลาไม่ได้ถูกปรับเปลี่ยนตามประสบการณ์ด้านอาหาร อยู่ภายใต้การควบคุมทางพันธุกรรมอย่างเข้มงวด และการถ่ายทอดทางพันธุกรรมของพวกมันเป็นไปตามธรรมชาติของคลินิก จากตัวอย่างของปลาหางนกยูงที่โตเต็มวัยทางเพศ พบว่าไม่มีความแตกต่างทางเพศในรสนิยมของปลา พบว่าความแตกต่างทางเพศสามารถแสดงออกมาได้ด้วยความเข้มข้นของการแสดงออกของปลาต่อพารามิเตอร์ต่างๆ ของการตอบสนองต่อรสชาติทางพฤติกรรม พบว่าปลาที่มีวิถีชีวิตต่างกันมีองค์ประกอบบางประการของการตอบสนองต่อรสชาติพฤติกรรมที่แตกต่างกัน (จำนวนการจับของเม็ด ระยะเวลาในการจับเม็ดในช่องปาก) พบว่าประสบการณ์การรับประทานอาหารที่แตกต่างกันไม่ส่งผลต่อรูปแบบการตอบสนองรสชาติโดยรวมของปลา
ผลการศึกษานี้สามารถใช้เป็นพื้นฐานสำหรับการพัฒนาและการแนะนำการเพาะเลี้ยงสัตว์น้ำโดยใช้สารกระตุ้นรสชาติทางเคมี ซึ่งออกแบบมาเพื่อเพิ่มประสิทธิภาพการกำหนดสูตรอาหารเทียมเพื่อเพิ่มความน่ารับประทานของปลาในฟาร์ม ผลลัพธ์ที่ได้สามารถนำไปใช้ในอุตสาหกรรมตู้ปลา (ปลาหางนกยูง, ปลาซิคลาโซมาเซเวอร์รัม) และการตกปลาเพื่อกีฬา (แมลงสาบ, ปลาคาร์พสีเงิน)
บทสรุป วิทยานิพนธ์ในหัวข้อ "Ichthyology", Nikolaeva, Ekaterina Valerievna
บทสรุป
ระบบรับสัมผัสทางเคมีหลักของปลา - การดมกลิ่นและการรับรส - สามารถมีส่วนร่วมในการก่อตัวของการตอบสนองเชิงพฤติกรรมของปลาต่อเม็ดที่มีสารเคมี การวิเคราะห์เปรียบเทียบการตอบสนองเชิงพฤติกรรมของปลาที่ไม่มีออสเมตและปลาควบคุม (ไม่บุบสลาย) (Kasumyan และ Morey, 1996) ซึ่งประเมินโดยใช้วิธีการที่เหมือนกับของเรา แสดงให้เห็นว่าปลาที่สูญเสียความไวในการรับกลิ่นไม่ได้นำไปสู่การเปลี่ยนแปลงปฏิกิริยาของพวกมันต่อเม็ดที่มี สาร ในปลาที่ไม่มีออสเมตและไม่บุบสลาย ความเข้มข้นเกณฑ์ของสารที่ใส่เข้าไปในแกรนูลก็ตรงกันเช่นกัน ข้อมูลเหล่านี้เป็นเหตุให้ยืนยันว่าระบบรับกลิ่นไม่เกี่ยวข้องกับการสนับสนุนทางประสาทสัมผัสในการตอบสนองเชิงพฤติกรรมของปลาทดลองที่เราสังเกตเห็นต่อเม็ดที่มีสารต่างๆ และธรรมชาติและความรุนแรงของการตอบสนองเหล่านี้ถูกควบคุมโดยการรับรู้รสชาติในช่องปาก
ผลการศึกษาพบว่าปลาที่มีระบบนิเวศต่างกันมีการรับรู้รสชาติที่ได้รับการพัฒนามาอย่างดี ทำให้สามารถแยกแยะคุณสมบัติรสชาติของสารประเภทต่างๆ ได้ การตอบสนองด้านรสชาติของปลานั้นถูกกำหนดโดยพันธุกรรมอย่างเคร่งครัด โดยในระหว่างการผสมข้ามพันธุ์ระหว่างปลาคาร์พ Silver Crucian และปลาคาร์พ พวกมันจะได้รับการสืบทอดตามประเภท Patroclinic ไม่มีความเชื่อมโยงระหว่างความชอบด้านรสชาติของปลากับสภาพที่อยู่อาศัยหรือรูปแบบการให้อาหารของปลา เห็นได้ชัดว่าเนื่องมาจากสเปกตรัมรสจำเพาะของสายพันธุ์ในระดับสูง ซึ่งแสดงออกมาไม่เพียงแต่ในด้านเคมีบำบัดเท่านั้น แต่ยังรวมถึงระดับสิ่งมีชีวิตด้วย คุณลักษณะนี้แสดงให้เห็นอย่างชัดเจนถึงความสำคัญและแน่นอนในการกำหนดบทบาทของการรับรู้รสชาติในการรับประกันการเลือกสรรโภชนาการของปลา รวมถึงความสามารถในการเลือกบริโภครายการอาหารที่เป็นลักษณะเฉพาะของพวกเขา
ข้อมูลที่ได้รับพร้อมกับข้อมูลที่มีอยู่เกี่ยวกับการไม่มีความจำเพาะของประชากรที่เห็นได้ชัดเจนในความชอบด้านรสชาติของปลา (Kasumyan, Sidorov, 1995) และการไม่มีอิทธิพลที่เห็นได้ชัดเจนของประสบการณ์การให้อาหารปลาแต่ละตัว (Kasumyan, Morey, 1997; Kasumyan , Morsy, 1998; ข้อมูลของเรา) ให้เหตุผลในการยืนยันว่าสเปกตรัมรสชาติในปลาถือได้ว่าเป็นลักษณะเฉพาะของจำนวนประชากรเฉพาะสายพันธุ์ โดยไม่ขึ้นกับเพศหรืออาหารที่บริโภค
รูปแบบการรับรู้รสชาติของปลาหลายรูปแบบและลักษณะเฉพาะของการตอบสนองต่อสารปรุงแต่งชนิดต่างๆ ถือเป็นประโยชน์ในทางปฏิบัติที่สำคัญ และสามารถนำมาใช้ในการแก้ปัญหาต่างๆ ในการประมงและการเพาะเลี้ยงสัตว์น้ำ การศึกษาที่ดำเนินการแสดงให้เห็นถึงคำมั่นสัญญาในการค้นหาและสร้างสารกระตุ้นและสารยับยั้งที่มีประสิทธิภาพสูงสำหรับปลา และทำหน้าที่เป็นพื้นฐานทางชีวภาพสำหรับการพัฒนาวิธีการควบคุมพฤติกรรมการให้อาหารปลาโดยใช้สิ่งกระตุ้นรสชาติ ผลลัพธ์ที่เราได้รับสามารถนำมาใช้เพื่อเพิ่มความอร่อยของอาหารสัตว์ ดำเนินการแก้ไขสูตรโดยการเติมสารพิเศษที่มีฤทธิ์กระตุ้นสูง หรือแยกส่วนประกอบที่มีสารประกอบยับยั้งออกจากอาหารสัตว์ สิ่งนี้จะไม่เพียงแต่ลดการสูญเสียอาหารเทียมโดยตรงในระหว่างการเพาะปลูกเท่านั้น แต่ยังช่วยให้มั่นใจในการเปลี่ยนอาหารสำหรับการเจริญเติบโตของปลาได้อย่างมีประสิทธิภาพมากขึ้น เนื่องจากเป็นที่ทราบกันดี (Takeda, Takii, 1992) ว่าการบริโภคอาหารที่มีคุณสมบัติดึงดูดใจทางเคมีนั้นมาพร้อมกับการหลั่งที่รุนแรงมากขึ้น ของเอนไซม์ย่อยอาหารในปลา
บรรณานุกรม วิทยานิพนธ์ทางชีววิทยา, ผู้สมัครสาขาวิทยาศาสตร์ชีวภาพ, Nikolaeva, Ekaterina Valerevna, มอสโก
1. อันดริยาเชฟ เอ.พี. พ.ศ. 2487 วิธีการรับอาหารจากทะเล ruffe Scorpaena porcus // Journal of General Biol. ต.5 ลำดับที่ 1 น.56-59.
2. อันดริยาเชฟ เอ.พี. พ.ศ. 24879 บทบาทของอวัยวะรับความรู้สึกในการหาอาหารในทะเล Burbot // Journal of General Biol. ต.5 ลำดับที่ 2. หน้า 123-127.
3. Arzhanova N.V., กรูเซวิช เอ.เค. พ.ศ. 2537 สภาวะอุทกเคมีในทะเลสีขาวในฤดูร้อนปี พ.ศ. 2534 // บทคัดย่อของ All-Russian การประชุม “ระบบนิเวศของทะเลรัสเซียภายใต้แรงกดดันจากมนุษย์ รวมถึงการประมง” แอสตราคาน 1994. หน้า 561-570.
4. Aristovskaya G.V. พ.ศ. 2478 ว่าด้วยเรื่องโภชนาการของปลาบางชนิด // Tr. แผนกตาตาร์ วนิโอรห์.
5. Aristovskaya G.V., Lukin A.V. พ.ศ. 2491 การประมงแม่น้ำสุระภายในสาธารณรัฐสังคมนิยมโซเวียตปกครองตนเองชูวัช // Tr. แผนกตาตาร์ วนิโอรห์. คาซาน.
6. อาร์โนลด์ ไอ.เอ็น. พ.ศ. 2445 การสังเกตโภชนาการของปลาในอ่างเก็บน้ำบางแห่งของ Valdai Upland // จากโรงงานปลา Nikolsky เอสพีบี ลำดับที่ 6.
7. บาร์ซูคอฟ วี.วี. พ.ศ. 2502 เสม. ปลาดุก (Anarchhididae) // สัตว์ในสหภาพโซเวียต ต.5 ฉบับที่ 5. ม.-ล. เอ็ด สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต
8. เบเรซินา เอ.พี. 2527. อุทกชีววิทยา. ม.: มัธยมปลาย. ป.203-204. โบโบรฟ ยู.พี. พ.ศ. 2511 โภชนาการและการเจริญเติบโตของปลาคาร์พหญ้าในสภาพฟาร์มบ่อในเขตภาคกลางของ RSFSR // การศึกษาใหม่เกี่ยวกับนิเวศวิทยาและการเพาะพันธุ์ปลาที่กินพืชเป็นอาหาร วิทยาศาสตร์. ม.106-115.
9. Bondarenko V.F. , Kuzmin S.Yu. 2527. การรับรู้กรดอะมิโนแต่ละตัวโดยปลาคาร์พอายุ 30 วัน // Min. ราศีมีน ครัวเรือนของสหภาพโซเวียต “ไบโอล. พื้นฐานของการเพาะเลี้ยงสัตว์น้ำอุตสาหกรรม” รวบรวมผลงานทางวิทยาศาสตร์เฉพาะเรื่อง คาลินินกราด. หน้า 78-91.
10. Bondarenko V.F., Kuzmin S.Yu., Shutov V.A. 2527. ปฏิกิริยาของตัวอ่อนปลาคาร์พต่อสารละลายกรดอะมิโนต่างๆ // การประมง. เลขที่ Yu.S. 30-31
11. Bondarenko V.F., Shutov V.A., Kuzmin S.Yu. 2529. การรับเคมีอาหารของตัวอ่อนปลาคาร์พ // ปลา. ครัวเรือน. ลำดับที่ 4. หน้า 33-34.
12. โบรุตสกี้ อี.วี. 2493 วัสดุเกี่ยวกับโภชนาการของปลาคาร์พ crucian เงิน (Carassius auratus gibelio Bloch) ในลุ่มน้ำอามูร์ // Tr. การสำรวจทางวิทยาอามูร์ ต. 1.
13. บรอนชไตน์ เอ.ไอ. 2493. รสชาติและกลิ่น. M.-JL: สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต 306 ส.
14. บรูวิช เอส.วี. พ.ศ. 2503 การศึกษาอุทกเคมีของทะเลสีขาว // Tr. สถาบัน สมุทรศาสตร์. ต. XLII หน้า 199-254.
15. บอแรกซ์ เอช.เจ. พ.ศ. 2516 ว่าด้วยโภชนาการของแมลงสาบ (Rutilus rutilus (L. ) ของ Curonian Lagoon // กระทรวงประมง เศรษฐกิจของสหภาพโซเวียต "การดำเนินการตามกฎหมาย" ฉบับ XLVI "ปัญหา Ichthyology" คาลินินกราด หน้า 37-43
16. วาซิเลฟสกายา เอ็น.อี. พ.ศ. 2517 ว่าด้วยการรับสารเคมีโดยไม่รับกลิ่นในปลา // ในคอลเลกชัน: ลักษณะพื้นฐานของพฤติกรรมและทิศทางของปลา อ.: วิทยาศาสตร์. หน้า 36-56.
17. Vasilevskaya N.E. , Nikitina I.P. พ.ศ. 2521 เครื่องมือตอบสนองแบบมีเงื่อนไขในปลาคาร์พต่อสิ่งเร้าเกลือและกรด // วารสาร สูงกว่า ประหม่า กิจกรรม ต. 28 ลำดับ 5 หน้า 1026-1036
18. Verigin B.V., เหงียนเวียด, เหงียนดง พ.ศ. 2506 วัสดุเกี่ยวกับการเลือกสรรอาหารและการปันส่วนรายวันของปลาคาร์พหญ้า // Mater ออล-ยูเนี่ยน สภา ตามฟาร์มเลี้ยงปลา การพัฒนาปลากินพืชเป็นอาหาร อาชกาบัต. 1963. หน้า 192-194.
19. โวเอโวดิน ดี.เอ็น. การให้อาหารลูกปลาในบริเวณชายฝั่งของช่องแคบ Great Salma // สื่อการประชุมทางวิทยาศาสตร์ของสถานีชีววิทยาทะเลสีขาวของมหาวิทยาลัยแห่งรัฐมอสโกซึ่งอุทิศให้กับความทรงจำของ N.A. Pertsov 17-18 สิงหาคม 2539 มอสโก หน้า 12-14.
20. วัชชานีนา แอล.ไอ. พ.ศ. 2515 อาหารแมลงสาบในอ่างเก็บน้ำคราเมนชูก // การประมง เคียฟ รีพับลิกัน
21. คอลเลกชันทางวิทยาศาสตร์เฉพาะเรื่องระหว่างแผนก ฉบับที่ 14. หน้า 91-97.
22. Gamygin E.A., Lysenko V.Ya., Sklyarov V.Ya., Turetsky V.I. 2532. อาหารผสมสำหรับปลา: การผลิตและวิธีการให้อาหาร. อ.: Agropromizdat. 168 น.
23. เกอร์ซา ไอ.ไอ. 2524. การส่องสว่างและพฤติกรรมของปลา. อ.: วิทยาศาสตร์. 163 หน้า แกรนด์ดิเลฟสกายา-เดกซ์บัค ม.ล. พ.ศ. 2504 คุณสมบัติหลักของสัตว์ด้านล่างและโภชนาการปลาของอ่างเก็บน้ำ Kama (พ.ศ. 2498-2502) // Tr. อูราล แผนก กอสNIORH. ลำดับที่ 5.
24. เกรซ ไอ.ไอ. พ.ศ. 2496 ในรูปแบบโภชนาการของแมลงสาบไซบีเรีย // Tr. รัฐทอมสค์ มหาวิทยาลัยที่ตั้งชื่อตาม V.V.Kuibysheva ตอมสค์ ต. 125.
25. Grigorash V.A., Spanovskaya V.D., Lebedeva T.V. พ.ศ. 2515 การจัดหาอาหารสำหรับลูกแมลงสาบ Rutilus rutilus (L. ) ในอ่างเก็บน้ำ Mozhaisk // Vopr. วิทยา ต. 12. ประเด็น 3 (74) หน้า 504-512.
26. Grigorash V.D., Spanovskaya, V.D., Lebedeva T.V. พ.ศ. 2516 การเจริญเติบโตและโภชนาการของแมลงสาบอายุน้อยในอ่างเก็บน้ำ Mozhaisk // มหาวิทยาลัยแห่งรัฐมอสโก "การศึกษาอ่างเก็บน้ำอย่างครอบคลุม" ฉบับที่ ครั้งที่สอง (เรียบเรียงโดย V.D. Bykov, N.Yu. Sokolova, K.K. Edelyptein) สำนักพิมพ์ของมหาวิทยาลัยแห่งรัฐมอสโก หน้า 166-18L
27. Devitsina G.V., คาซเลฟ เอ.เอ. พ.ศ. 2538 ระบบเคมีเซ็นเซอร์และการสร้างสัณฐานวิทยาแบบเฮเทอโรโครนิกในปลาสเตอร์เจียนรุ่นเยาว์ตอนต้น // ชีวฟิสิกส์ ต. 40. ลำดับ 1. หน้า 146-150
28. Denisov, Dzhenyuk, 1995. ทรัพยากรชีวภาพของเขตชายฝั่งทะเลของคาบสมุทร Kola // สมัยใหม่ สภาพและการใช้งานอย่างมีเหตุผล ไม่แยแส หน้า 10-25.
29. Dmitrieva E.N. 2500 การวิเคราะห์ทางสัณฐานวิทยาของปลาคาร์พ crucian สองสายพันธุ์ // Tr. สถาบันสัณฐานวิทยาสัตว์ตั้งชื่อตาม เอ.เอ็น. เซเวิร์ตโซวา ต.16.
30. ดอมราเชฟ พี.เอฟ., ปราฟดิน ไอ.เอฟ. พ.ศ. 2505 ปลาในทะเลสาบอิลเมนและแม่น้ำ Volkhov และความสำคัญด้านการประมง // Mat. จากการวิจัยพบว่า R. Volkhov และแอ่งน้ำ เลนินกราด ฉบับที่ X
31. ดราเชฟ เอส.เอ็ม. พ.ศ. 2511 การเปลี่ยนแปลงองค์ประกอบทางเคมีและคุณสมบัติของน้ำในแม่น้ำมอสโกเนื่องจากมลภาวะและกระบวนการทำให้บริสุทธิ์ในตัวเอง // TR. อินทาไบโอล. นานาชาติ น่านน้ำของสหภาพโซเวียต “เคมีของแหล่งน้ำภายในประเทศและปัจจัยของมลพิษและการทำให้บริสุทธิ์ในตัวเอง” ต.18 (2) หน้า 152-198.
32. เอลิซอฟ วี.วี. 2540. การประเมินความร้อนของน้ำและความสมดุลของเกลือของทะเลสีขาว // “ อุตุนิยมวิทยาและอุทกวิทยา”. ลำดับที่ 9. หน้า 83-93.
33. เออร์โมลิน วี.พี. พ.ศ. 2520 การให้อาหารทรายแดง ปลาสเตอร์เล็ต ทรายแดงเงิน และแมลงสาบในอ่างเก็บน้ำ Saratov // Tr. แผนก Sarotovsky กอสนิออร์คา. ซาราตอฟ. ต. XV. "โวลก้าตอนล่างและอ่างเก็บน้ำ" หน้า 75-78.
34. เชลเทนโควา เอ็ม.วี. พ.ศ. 2492 องค์ประกอบอาหารและการเจริญเติบโตของตัวแทนสกุล Putilus // Zool นิตยสาร ต. 28. ประเด็น. 3.
35. เชลเทนโควา เอ็ม.วี. พ.ศ. 2494 เรื่องความเป็นพลาสติกในอาหารของแมลงสาบ // Tr. มีสติทั้งหมด การวิจัยทางวิทยาศาสตร์ สถาบันประมงทะเล. เศรษฐศาสตร์และสมุทรศาสตร์ (VNIRO) ต.18.
36. เชลเทนโควา เอ็ม.วี. พ.ศ. 2503 ว่าด้วยความสัมพันธ์ทางอาหารของแมลงสาบและทรายแดงในทะเลสาบ Curonian // Tr. มีสติทั้งหมด การวิจัยทางวิทยาศาสตร์ ปลาอินทา เศรษฐศาสตร์และสมุทรศาสตร์ (VNIRO) ต.42.
37. อิวาโนวา เอ็น.ที. พ.ศ. 2498 ความสำคัญทางชีววิทยาและการประมงของปลาคาร์พ crucian เงินในอ่างเก็บน้ำ Veselovsky // บันทึกทางวิทยาศาสตร์ของมหาวิทยาลัย Rostov-on-Don ต.29.
38. อิฟเลฟ VS. 2520. ทดลองนิเวศวิทยาการให้อาหารปลา. เคียฟ: Nauk.dumka. 272 หน้า
39. อิลยิน เอ็ม.เอ็น. 2508. การเลี้ยงปลาในตู้ปลา. อ.: มหาวิทยาลัยแห่งรัฐมอสโก. หน้า 248-249.
40. อิลยิน เอ็ม.เอ็น. 2511. การเลี้ยงปลาในตู้ปลา. อ.: สำนักพิมพ์มหาวิทยาลัยมอสโก. 399 หน้า
41. คาสซิล วี.จี. พ.ศ. 2515 รสชาติ // สรีรวิทยาของระบบประสาทสัมผัส. 4.2. ล.: วิทยาศาสตร์. หน้า 562-606.
42. คาสซิล วี.จี. 2533. พฤติกรรมการกินในการถ่ายทอดทางพันธุกรรม. ล.: วิทยาศาสตร์. 220 หน้า
43. Kasumyan A.O., Pashchenko N.I. พ.ศ. 2525 การประเมินบทบาทของกลิ่นในปฏิกิริยาป้องกันของปลาคาร์พหญ้า Ctenopharyngodon idella (Val.) (Cyprinidae) ต่อฟีโรโมนปลุก // ปัญหาของวิทยาวิทยา ต. 22. ประเด็น 2. หน้า 303-307.
44. Kasumyan A.O., Sidorov S.S., Pashchenko N.I., Nemchinov A.V. 2535 ความไวต่อรสชาติภายนอกและในช่องปากของปลาสเตอร์เจียนรัสเซียวัยเยาว์ Acipenser gueldenstaedti ต่อกรดอะมิโน // Dokl. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต. 322 ลำดับ 1 หน้า 193-195
45. คำถามเกี่ยวกับวิทยาวิทยา ต. 34 ลำดับ 6 หน้า 831-838 พ.ศ. 2538 การวิเคราะห์เปรียบเทียบการตอบสนองด้านรสชาติของปลาเทราต์เด็กและเยาวชน Salmo trutta trutta จากประชากรในทะเลบอลติกและทะเลสีขาว // รายงานของ Academy of Sciences ต. 343 ลำดับ 3 หน้า 417-419
46. Kasumyan A.O., Sidorov S.S., Pashchenko N.I. 1993ก. อิทธิพลของอุณหภูมิของน้ำต่อความไวต่อรสชาติของปลาสเตอร์เจียนสเตลเลตวัยเยาว์ Acipenser stellatus ต่อกรดอะมิโนอิสระ // ดอกกล. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต. 331 ลำดับที่ 2 หน้า 248-250.
47. คาสุมยาน เอ.โอ., โมเรย์ เอ.เอ็ม.ค., ซิโดรอฟ เอส.เอส. พ.ศ. 2536. ความไวต่อรสชาติของปลาคาร์พ Cyprynus carpio ต่อสารที่ทำให้เกิดความรู้สึกรับรสประเภทหลัก // Dokl. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต.330. ลำดับที่ 6.ส. 792-793.
48. เกษมยาน A.O., Taufik J.P. 2536. พฤติกรรมตอบสนองของปลาสเตอร์เจียนวัยอ่อน (Acipenseridae) ต่อกรดอะมิโน // Vopr. วิทยา ต. 33. ประเด็น. 5. หน้า 691-700.
49. เกษมยาน อ.โอ., เดวิทสินา จี.วี. พ.ศ. 2540 อิทธิพลของการกีดกันดมกลิ่นต่อความไวของสารเคมีและสถานะของต่อมรับรสของปลาสเตอร์เจียน // ประเด็น. วิทยา ต. 37 ลำดับ 6 หน้า 823-835
50. Kasumyan A.O., Nikolaeva E.V. พ.ศ. 2540 ความชอบด้านรสชาติของปลาหางนกยูง Poecilia reticulata (Cyprinodontiformes) // ปัญหา วิทยา ต. 37 ลำดับ 5 หน้า 696-703
51. Katasonov V.Ya., Gomelsky B.I. พ.ศ. 2534 การคัดเลือกปลาที่มีพื้นฐานทางพันธุศาสตร์ M.: VO "Agropromizdat" ป.208.
52. เคอร์พิชนิคอฟ VS. พ.ศ. 2522 พื้นฐานทางพันธุกรรมของการคัดเลือกปลา // เลนินกราด วิทยาศาสตร์. เลนินกรา. แผนก 392 หน้า
53. กลูชาเรวา โอ.เอ. พ.ศ. 2503 การให้อาหารปลากินเนื้อในอ่างเก็บน้ำ Rybinsk // Tr. ดาร์วิน, แซป. ที่อ่างเก็บน้ำ Rybinsk ม.ฉบับ. 6.
54. โครเกียส เอฟ.วี., โครคิน อี.เอ็ม., เมนชัตกิน วี.วี. 2512 ชุมชนปลาผิวน้ำในทะเลสาบ Dalney (ประสบการณ์การสร้างแบบจำลองไซเบอร์เนติกส์) L.: Nauka 86ส
55. Kublickas A. 1959. การให้อาหารปลาหน้าดินในอ่าว Kurplo Mares. วิลนีอุส SSR วรรณกรรม
56. คุดรินสกายา โอ.เอ. 2509. ในประเด็นโภชนาการของปลาไพค์คอน ทรายแดง และแมลงสาบ // วารสารทางอุทกชีววิทยา. ต.2. ฉบับที่ 6. /
57. Lebedev V.D., Spanovskaya V.D. พ.ศ. 2526 ครอบครัว Cyprinidae // M. ชีวิตสัตว์. ต.4. ปลา. ป.228-272.
58. ลูปาเชวา แอล.ไอ. พ.ศ. 2510 การให้อาหารปลาคาร์พหญ้าในระยะแรกของการแตกหัก // Rybn. ครัวเรือน เคียฟ ฉบับที่ 3. หน้า 102-104.
59. Maksimenkov V.V. , Tokranov A.M. พ.ศ. 2536 การให้อาหาร Hypomesus olidus จากปากเล็กที่ปากแม่น้ำ Bolypaya (Western Kamchatka) // Vopr. วิทยา ต. 33 ลำดับ 3 หน้า 388-394
60. Maksimenkov V.V. , Tokranov A.M. พ.ศ. 2537 การให้อาหาร Pungitius pungitius (Gasterosteidae) หนามเก้าหนามในบริเวณปากแม่น้ำและตอนล่างของแม่น้ำ Bolshaya (Western Kamchatka) // Vopr. วิทยา ต. 34 ลำดับ 5 หน้า 697-702
61. Maksimova M.P. , Nesvetova G.I. , Chugainova V.A. พ.ศ. 2532 สภาวะอุทกเคมีในน่านน้ำของอ่าว Kandalaksha และ Onega ของทะเลสีขาว // Coll. ทางวิทยาศาสตร์ การดำเนินการ “ปัญหาสมุทรศาสตร์เชิงพาณิชย์ลุ่มน้ำภาคเหนือ” มูร์มันสค์. หน้า 166-176.
62. มายูคินา จี.เอ., ดมิตรีวา เอ็น.จี., มารุซอฟ อี.เอ., ยัวร์เควิช จี.วี. พ.ศ. 2512 การดมกลิ่นและบทบาทต่อพฤติกรรมของปลา // ผลลัพธ์ของวิทยาศาสตร์. 2511. สัตววิทยา. อ.: สำนักพิมพ์ VINITI AN USSR ป.32-78.
63. Manteifel B.P., Girsa I.I., Leshcheva T.S., Pavlov D.S. พ.ศ. 2508 จังหวะโภชนาการและการเคลื่อนไหวของปลานักล่าในน้ำจืดบางชนิดในแต่ละวัน // ในคอลเลกชัน: โภชนาการของปลานักล่าและความสัมพันธ์กับสิ่งมีชีวิตในอาหาร อ.: วิทยาศาสตร์ ป.3-81.
64. เมยุค เอ็น.เอส. พ.ศ. 2498 โภชนาการของปลามูลค่าต่ำและเป็นวัชพืชของ Lower Dnieper // Tr. วิจัย สถาบันประมงบ่อน้ำและทะเลสาบ-แม่น้ำ ครัวเรือน เคียฟ ลำดับที่ 10.
65. เนโบลซินา ที.เค. พ.ศ. 2508 การประเมินคุณภาพและเชิงปริมาณของโภชนาการของทรายแดง ทรายแดงเงิน และแมลงสาบในอ่างเก็บน้ำโวลโกกราด พ.ศ. 2505-2507 //ต.อ.ซาราตอฟ สถาบันวิจัยและพัฒนาแห่งรัฐ ซาราตอฟ. ต.8
66. Nikolaeva E.V., Kasumyan A.O. 2543. การวิเคราะห์เปรียบเทียบความชอบด้านรสชาติและการตอบสนองเชิงพฤติกรรมต่อสิ่งเร้ารสชาติในปลาหางนกยูงตัวเมียและตัวผู้ Poecilia reticulata // ฉบับที่ วิทยา ต.40. ลำดับที่ 4. หน้า 560-565.
67. Nikolsky G.V. , Sokolova N.Yu. พ.ศ. 2493 เกี่ยวกับวิธีการประมง การพัฒนาแหล่งน้ำในพื้นที่พักพิงป่า Kamyshin-Volgograd // “ สวนสัตว์ วารสาร." ต. XXXX ฉบับที่ 3.
68. นิโคลสกี้ จี.วี. 2517. นิเวศวิทยาของปลา. ม.: มัธยมปลาย. 174 ส.
69. พาฟโลฟ ดี.เอส., คาสุมยาน เอ.โอ. 2533. ฐานประสาทสัมผัสของพฤติกรรมการให้อาหารปลา // ปัญหาวิทยาวิทยา. 1990. ที.โซ. ฉบับที่ 5. ป.720-732.
70. พาฟโลฟ ดี.เอส., คาสุมยาน เอ.โอ. 2541. โครงสร้างพฤติกรรมการให้อาหารปลา // ประเด็นวิทยาวิทยา. 2541 ต.38. ลำดับที่ 1 หน้า 123-136
71. Pevzner R. A. 1980. คุณสมบัติวิวัฒนาการบางประการของการจัดระเบียบอวัยวะรับรสของปลา // ในการรวบรวม "ระบบประสาทสัมผัส กลิ่นและรส ชล: วิทยาศาสตร์ หน้า 82-93
72. เพฟซเนอร์ อาร์.เอ. พ.ศ. 2527. บทบาทของระบบประสาทสัมผัสต่อพฤติกรรมการให้อาหารของปลาสเตอร์เจียน // รวบรวม. : สรีรวิทยาทางประสาทสัมผัสของปลา ไม่แยแส ป.36.
73. เพฟซเนอร์ อาร์.เอ. 2528. โครงสร้างการจัดโครงสร้างของตัวรับรสในปลากระดูกพรุน. สาม. ตัวอ่อนในช่วงให้นมไข่แดง // Cytology. ต.27. ลำดับที่ 11 หน้า 1240-1246
74. Petrovitsky I. 1984. พิพิธภัณฑ์สัตว์น้ำปลาเขตร้อน อาร์เทีย. ปราก 224ส.
75. พินสกี เอฟ.ยา. 2510. การให้อาหารปลาสติ๊กเกิลแบ็ก (Pungitius pungitius L.) และความสัมพันธ์ทางอาหารของมันกับปลาแซลมอนวัยอ่อน (Salmo salar L.) ในบ่อของโรงเพาะฟักปลา Tome ใน Latvian SSR. Izv. สถาบันวิจัยประมงทะเลสาบและแม่น้ำ ครัวเรือน ต. 63. หน้า 202-205.
76. พอดดับนี เอ.จี. พ.ศ. 2509 ในการตอบสนองแบบปรับตัวของประชากรแมลงสาบต่อการเปลี่ยนแปลงสภาพที่อยู่อาศัย // Tr. สถาบันไบโอล น้ำภายใน สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ฉบับที่ 10 (13)
77. โปลิวานนายา M.F., วัชชานีนา แอล.ไอ. พ.ศ. 2512 การให้อาหารแมลงสาบในอ่างเก็บน้ำคราเมนชูก // Coll. "ประมง" เค "เก็บเกี่ยว" ฉบับที่ 8.
78. โปลอนสกี้ เอ.เอส. 1991ก. การเก็บรักษาและเพาะพันธุ์ปลาในตู้ปลา มอสโก VO "Agropromizdat" 384 หน้า
79. โปลอนสกี้ เอ.เอส. พ.ศ. 2534 การเพาะพันธุ์ปลาหางนกยูงในสิงคโปร์ // การประมง ลำดับที่ 10 น.82-85.
80. โปปอฟ ป.เอ. พ.ศ. 2518 ลักษณะทางสัณฐานวิทยาของไซบีเรียนเดซ Leuciscus leuciscus baicalensis (Dyb.) พูลพี ธนาม // คำถาม. วิทยา ต. 15. ประเด็น 6 (95) หน้า 1011-1012.
81. ปุชคอฟ เอ็น.วี. 2497 สรีรวิทยาของปลา // Pishchepromizdat. ม.
82. โรดิโอโนวา แอล.เอ. พ.ศ. 2512 การให้อาหารแมลงสาบในอ่างเก็บน้ำ Kama // บันทึกทางวิทยาศาสตร์ของมหาวิทยาลัยระดับการใช้งาน “ วัสดุของการประชุมทางวิทยาศาสตร์และการผลิตเกี่ยวกับสถานะของฐานวัตถุดิบของอ่างเก็บน้ำของภูมิภาคระดับการใช้งานและการใช้ประโยชน์ประมง” เพอร์เมียน
83. ซาบาเนฟ แอล.พี. พ.ศ. 2454 ปลาแห่งรัสเซีย ชีวิตและการตกปลา (ตกปลา) ของปลาน้ำจืดของเรา M. จัดพิมพ์โดย A.A. Kartsev 1,062 หน้า
84. ซาวินา เอ็น.โอ. พ.ศ. 2501 ปลาคาร์พสีเงิน Crucian Carassius auratus gibelio (Bloch) ในสภาพที่อยู่อาศัยใหม่ // Tr. สถาบันประมงเบลารุส ต.11.
85. สแปนอฟสกายา วี.ดี. พ.ศ. 2491 การให้อาหารปลาในอ่างเก็บน้ำ Uchinsky // Zool นิตยสาร ต.27. ฉบับที่ 1.
86. สเตปาโนวา. บน. พ.ศ. 2496 การใช้ประมงในอ่างเก็บน้ำ Katta-Kurgan // Izv. สถาบันวิทยาศาสตร์แห่ง UzSSR ฉบับที่ 5.
87. สทิจ ที.เอส. พ.ศ. 2516 การศึกษาทดลองการให้อาหารปลาคาร์พหญ้าวัยอ่อน // Kazakh Academy of Sciences สสส. นิเวศวิทยาของไฮโดรไบโอออนต์ในแหล่งน้ำของคาซัคสถาน อัลมา-อาตา. 136-142.
88. ซูโฮเวอร์คอฟ เอฟ.เอ็ม. 2494. ความสำคัญทางเศรษฐกิจของปลาคาร์พสีเงินในการเลี้ยงปลาในบ่อ // “การประมง”. ลำดับที่ 4.
89. Tamar G. 1976. พื้นฐานของสรีรวิทยาทางประสาทสัมผัส. ม. มีร์. 520 หน้า
90. Tokranov A.M. , Maksimenkov V.V. พ.ศ. 2536 ลักษณะการกินอาหารของปลาลิ้นหมาดาวปลาลิ้นหมา Platichthys stellatus ในบริเวณปากแม่น้ำ Bolshaya (คัมชัตกาตะวันตก) // ฉบับที่. วิทยา ต.33.ลำดับ4.ป.561-565.
91. เคเรมิโซวา เค.เอ. พ.ศ. 2501 โภชนาการของปลาไวท์ฟิชและปลาคาร์พสีเงินในแหล่งที่อยู่อาศัยใหม่ // Tr. สถาบันอุทกชีววิทยาเบลารุส ต.11.
92. เชอร์ฟาส เอ็น.บี., อับราเมนโก เอ็ม.เอ็น., เอเมลยาโนวา โอ.วี., อิลิน่า ไอ.ดี., ทรูเวลเลอร์ เค.เอ. 2529. ลักษณะทางพันธุกรรมของการเกิดจีโนเจเนซิสในลูกผสมของปลาคาร์พ crucian เงินและปลาคาร์พ // พันธุศาสตร์. ต.22.ข้อ 1.หน้า134-139.
93. Harborne J. 1985. ชีวเคมีสิ่งแวดล้อมเบื้องต้น. ม. มีร์. 312 หน้า
94. คาริโตโนวา เอ็น.เอ็น. 2506. ความสัมพันธ์ทางโภชนาการและอาหารของปลาคาร์พและปลาคาร์พสีเงินในบ่อ // Ukr. สถาบันวิจัยปลา. ครัวเรือน. "การเพิ่มผลผลิตปลาในบ่อ" ทางวิทยาศาสตร์ ต. ต. XV. หน้า 7-25.
95. คเลโบวิช วี.วี. พ.ศ. 2505 คุณสมบัติขององค์ประกอบของสัตว์น้ำขึ้นอยู่กับความเค็มของสิ่งแวดล้อม // Zhurn ทั่วไป ไบโอล ต. XXIII ลำดับที่ 2. หน้า 90-97.
96. คเลโบวิช วี.วี. พ.ศ. 2517 ความเค็มวิกฤตของกระบวนการทางชีววิทยา // “ วิทยาศาสตร์” เลนินกราด 236 หน้า
97. โคมเชนโก ไอ.จี. พ.ศ. 2534 การเปรียบเทียบองค์ประกอบไฮโดรเคมีของน้ำในแหล่งเก็บน้ำของประเทศเขตร้อนและสหภาพโซเวียต // "การประมง" ลำดับที่ 6 หน้า 9294
98. Fedorova" G.V., Tikhomirova L.P. 1980. การให้อาหารแมลงสาบในทะเลสาบ Ladoga // การรวบรวมผลงานทางวิทยาศาสตร์ ฉบับที่ 159. "ชีววิทยาและการประมงของทะเลสาบขนาดใหญ่ทางตะวันตกเฉียงเหนือ" (Ed. G. V. Fedorova, Leningrad , หน้า 31-41.
99. แฟรงค์ เซนต์. พ.ศ. 2518 สารานุกรมภาพประกอบของปลา // อาร์เทีย ป.432.
100. เซปคิน อี.เอ., โซโคลอฟ แอล.ไอ. พ.ศ. 2541 ในการแพร่กระจายอย่างเข้มข้นของ Pungitius pungitius (L. ) ที่มีหนามเก้าหนามในลุ่มน้ำมอสโก // Vestn มหาวิทยาลัยแห่งรัฐมอสโก เซอร์ 16. ชีววิทยา. ลำดับที่ 3. หน้า 37-39.
101. ชามาร์ดินา ไอ.พี. พ.ศ. 2514 การให้อาหารแมลงสาบในอ่างเก็บน้ำ Mozhaisk // มหาวิทยาลัยแห่งรัฐมอสโก “ การศึกษาอ่างเก็บน้ำอย่างครอบคลุม” ฉบับที่ 1. (เรียบเรียงโดย V.D. Bykov) ป.250.
102. ชิโวเคเน่ Y.S. 2526 องค์ประกอบเชิงคุณภาพและเชิงปริมาณของกรดอะมิโนที่ถูกผูกมัดและอิสระในอาหารธรรมชาติและอาหารสำหรับปลาในบ่อ // Tr. SSR วรรณกรรม เซอร์.วี. ต.3(83) ป.81-85.
103. ชมิดตอฟ เอ.ไอ. พ.ศ. 2495 บทบาทของแมลงสาบในการประมงของ Tatar AS และลักษณะทางชีววิทยาใน Kama ตอนล่างและแม่น้ำโวลก้าตอนกลาง // Uch. แซบ มหาวิทยาลัยคาซาน. คาซาน. ต. 112. หนังสือ. 7.
104. Shpet G.I., Kharitonova N.N., Bakunenko L.A. พ.ศ. 2493 ว่าด้วยการเปรียบเทียบสัณฐานวิทยาของอุปกรณ์เหงือกของปลาคาร์พ crucian และปลาคาร์พ crucian ที่เกี่ยวข้องกับความแตกต่างทางโภชนาการ // Zool นิตยสาร." ที.เอ็กซ์แอล. ฉบับที่ สิบเอ็ด
105. Shubnikov D.A., Pshenichny B.P., Chestnova L.G. พ.ศ. 2513 วัสดุทางชีววิทยาของปลาลิ้นหมาขั้วโลกในอ่าว Kandalaksha ของทะเลสีขาว // ใน: ชีววิทยาของทะเลสีขาว การดำเนินการของสถานีชีววิทยาทะเลสีขาวของมหาวิทยาลัยแห่งรัฐมอสโก ที.ซี. สำนักพิมพ์ของมหาวิทยาลัยแห่งรัฐมอสโก หน้า 189-207.
106. ยาโรเชนโก M.F. , Naberezhny A.N. , Valkovskaya O.I. พ.ศ. 2496 ความสัมพันธ์ทางอาหารระหว่างปลาคาร์พกับปลาคาร์พ crucian เมื่อปลูกร่วมกันในบ่อในมอลโดวา // ข่าวสาขามอลโดวาของสถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ฉบับที่ 5.
107. Aburto-Oropeza O., Sala E., Sanchez-Ortiz S. 2000. พฤติกรรมการให้อาหาร, การใช้แหล่งที่อยู่อาศัย, และความอุดมสมบูรณ์ของปลาเทวดา Holacanthus passer (Pomacanthidae) ในทะเลคอร์เตสตอนใต้ // ชีววิทยาสิ่งแวดล้อมของปลา. ว. 57. หน้า 435-442.
108. Adams M.A., Johnsen P.B., Hong-Qi Z. 1988. การเพิ่มประสิทธิภาพทางเคมีในการให้อาหารสำหรับปลาที่กินพืชเป็นอาหาร Tilapia zillii // การเพาะเลี้ยงสัตว์น้ำ. U.12 เลขที่ 12. หน้า 95-107.
109. Appelbaum S. 1980. Veruche zur Geschmacksperzeption einiger SusSwasserfische im larvalen und adulten Stadium // Arch. ฟิชเชอไรวิส. บด.31 เลขที่ 2. หน้า 105-114.
110. Appelbaum S., Adron J.W., George S.G., Mackie A.M., B.J.S.Pirie 2526. พัฒนาการของการดมกลิ่นและอวัยวะรับรสของ Solea solea ระหว่างการเปลี่ยนแปลง // เจ.มี.ค. ไบโอล ตูด สหราชอาณาจักร ว. 63>. 97-108.
111. Atema J. 1971. โครงสร้างและหน้าที่ของความรู้สึกรับรสในปลาดุก (Ictalurus natalis) // สมอง, พฤติกรรมและวิวัฒนาการ. โวลต์ 4. หน้า 273-294.
112. Atema J. 1980. ความรู้สึกทางเคมี สัญญาณทางเคมี และพฤติกรรมการให้อาหารในปลา // พฤติกรรมของปลาและการใช้ในการจับและเพาะเลี้ยงปลา มะนิลา. ป.57-101.
113. Adron J.W., Mackie A.M. 2521 การศึกษาลักษณะทางเคมีของการให้อาหารสารกระตุ้นสำหรับเรนโบว์เทราท์ Salmo gairdneri Richardson // J. Fish Biol ว.12. ลำดับที่. 4. หน้า 303-310.
114. เบิร์ด อาร์.ซี. 2508 ผลกระทบทางนิเวศวิทยาของพฤติกรรมของปลาบู่ Dimorphic ทางเพศ Microgobius gulosus (Girard) // Publ. สถาบัน วิทยาศาสตร์ทางทะเล เท็กซัส ว. 10. ป. 1-8.
115. แบมส์ อาร์.เอ. พ.ศ. 2519 การอยู่รอดและแนวโน้มในการกลับบ้านซึ่งได้รับผลกระทบจากการมีหรือไม่มียีนของบิดาที่ดัดแปลงในท้องถิ่นในประชากรปลาแซลมอนสีชมพูที่ปลูกถ่ายสองกลุ่ม (Oncorhynchus gorbuscha) // J. Fish ความละเอียด บี.แคนาดา. ว.33.ลำดับที่. 12. หน้า 2716-2725.
116. บาร์ดาค เจ.อี., วินน์ เอช.อี., เมนเซล ดี.ดับเบิลยู. พ.ศ. 2502 บทบาทของประสาทสัมผัสในการให้อาหารแก่นักล่าแนวปะการังในเวลากลางคืน Gymnothorax moringa และ G. vicinus // Copeia เลขที่ 2. หน้า 133-139.
117. บาร์ดาช เจ.อี., ท็อดด์ เจ.เอช., คริกเมอร์ อาร์.เค. 2510. จำแนกตามรสชาติของปลาสกุล Ictalurus. // วิทยาศาสตร์, V. 155, P. 1276-1278.
118. แบร์ติโน เอ็ม., โบชอมป์ จี.เค., เจน เค.ซี. พ.ศ. 2526 จัดอันดับการรับรู้รสชาติในสองกลุ่มวัฒนธรรม // ความรู้สึกทางเคมี V.8 หมายเลข 1 หน้า 3-15
119. แบรนสัน วี.เอ. พ.ศ. 2509 การสังเกตทางเนื้อเยื่อวิทยาของปลาสเตอร์เจียนปลาสเตอร์เจียน Hybopsis gelida (Cyprinidae) // Copeila เลขที่ 4. หน้า 872-876.
120. Breder C.M., Rasquin P. 2486. ปฏิกิริยาทางประสาทสัมผัสทางเคมีใน characins คนตาบอดชาวเม็กซิกัน // Zoologica. ว.28.ป.169-200.
121.ธ. บุษนิตา ศรี เอ.คริสเตียน. 2502. Biologia si cresterea carasului argintiu (Carassius auratus gibelio (Bloch)) ใน iazuri si elesteele de crap. Buletenul institutului de cercetari piscicole Anul. 18.เลขที่ 1.
122. คาเมรอน เจ.เอ็น., คอสโตริส เจ., เพนฮาเล พี.เอ. พ.ศ. 2516 งบประมาณพลังงานเบื้องต้นของ Pungitius pungitius เก้ากระดูกสันหลังในทะเลสาบ Arctc //
123. เจฟิช ความละเอียด บด. แคนาดา 30. หน้า 1179-1189
124. Caprio J. 1975. ความไวสูงของตัวรับรสปลาดุกต่อกรดอะมิโน // Sotr. ไบโอเคม ฟิสิออล. ว. 52เอ. ป.247-251.
125. Caprio J. 1978. การดมกลิ่นและรสชาติในปลาดุกแชนเนล: การศึกษาทางไฟฟ้าสรีรวิทยาของการตอบสนองต่อกรดอะมิโนและอนุพันธ์ // J. Sotr. ฟิสิออล. ว. 123. หน้า 357-371.
126. Caprio J. 1982. ความไวและความจำเพาะสูงของตัวรับกลิ่นและการรับรสของปลาดุกต่อกรดอะมิโน // ใน: Chemoreception in-Fishes (Ed. T. J. Hara) อัมสเตอร์ดัม: Elsevier Scientific Publishing Al. ป.109-134.
127. Caprio J. 1984. การดมกลิ่นและรสชาติในปลา // ใน: สรีรวิทยาเปรียบเทียบของระบบประสาทสัมผัส (Eds. L. Bolis, R. D. Keynes และ S. H. P. Maddrell) มหาวิทยาลัยเคมบริดจ์ กด. ป.257-283.
128. Caprio J. 1988. ตัวกรองอุปกรณ์ต่อพ่วงและเซลล์รับเคมีบำบัดในปลา // ใน: ชีววิทยาทางประสาทสัมผัสของสัตว์น้ำ (Ed. J. Atema, R. R. Fay, A. N. Popper, W. N. Tawolga) สปริงเกอร์-แวร์แลก เบอร์ลิน ป.313-338.
129. คาร์ W.E.S. 2525 การกระตุ้นทางเคมีของพฤติกรรมการให้อาหาร // ใน: Chemoreception ในปลา (Ed. T. J. Hara) อัมสเตอร์ดัม: Elsevier Scientific Publishing Al. ป.259-273.
130. Dabrowski JC., Rusiecki M. 1983. เนื้อหาของกรดอะมิโนทั้งหมดและอิสระในอาหารแพลงก์ตอนสัตว์ของตัวอ่อนปลา // การเพาะเลี้ยงสัตว์น้ำ. ว.30.ลำดับที่. 1-4. ป.31-42.
131. Davenport C.J., Caprio J. 1982. การบันทึกรสชาติและการสัมผัสจาก ramus เกิดขึ้นซ้ำบนใบหน้าทำให้ต่อมรับรสด้านข้างในปลาดุก // J. Sotr ฟิสิออล. ก.V. 147. หน้า 217-229.
132. De la Noue J., Choubert G. 1985. การย่อยได้ชัดเจนของชีวมวลที่ไม่มีกระดูกสันหลังด้วยเรนโบว์เทราต์ // การเพาะเลี้ยงสัตว์น้ำ. ว. 50. หน้า 103-112.
133. ดัสซอลท์ จี.วี., เครเมอร์ ดี.แอล. พ.ศ. 2524 พฤติกรรมการกินอาหารและการกินอาหารของปลาหางนกยูง Poecilia reticulata (Pisces: Poeciliidae) // Can. เจ.ซูล. ว. 59. ลำดับที่. 4. หน้า 684-701.
134. Farr J.A., Herrnkind W.F. พ.ศ. 2517 การวิเคราะห์เชิงปริมาณของปฏิสัมพันธ์ทางสังคมของปลาหางนกยูง Poecilia reticulata (Pisces: Poeciliidae) เป็นฟังก์ชันของความหนาแน่นของประชากร // Anim ประพฤติตน ว. 22. หน้า 582-591.
135. ฟาร์ เจ.เอ. พ.ศ. 2518 บทบาทของการปล้นสะดมในวิวัฒนาการของพฤติกรรมทางสังคมของประชากรตามธรรมชาติของปลาหางนกยูง Poecilia reticulata (Pisces: Poeciliidae) // วิวัฒนาการ ว.29.ป.151-158.
136. Finger T.E., Morita Y. 2528. ระบบรับรสสองระบบ: นิวเคลียสรับรสของใบหน้าและช่องคลอดมีการเชื่อมต่อก้านสมองที่แตกต่างกัน // วิทยาศาสตร์ โวลต์ 227. หน้า 776-778.
137. Finger T.E., Drake S.K., Kotrschal K., Womble M., Dockstader K.C. 2534. การเจริญเติบโตหลังตัวอ่อนของระบบรับรสส่วนปลายในช่องปลาดุก, Ictalurus punctatus // J. Compar. ประสาทวิทยา. ว. 314. หน้า 55-66.
138. Gerhart D.J., Bondura M.E., Commito J.A. พ.ศ. 2534 การยับยั้งการให้อาหารปลาซันฟิชโดยสเตียรอยด์ป้องกันจากแมลงเต่าทอง: ความสัมพันธ์ระหว่างโครงสร้างกับกิจกรรม // J. Chem อีคอล. ว.17.ลำดับที่. 7. หน้า 1363-1370.
139. Glaser D. 1966. Unterschungen uber ตายแน่นอน Geschmacksschwellen von Fischen // Z. Vergl ฟิสิออล. บด. 52. ป.1-25.
140. Goh Y., Tamura T. 1980a. การตอบสนองทางกลิ่นและการรับรสต่อกรดอะมิโนในลมหายใจจากทะเลแดงและปลากระบอก 2 ชนิด // Sotr. ไบโอเคม ฟิสิออล. วี 66ซี ป.217-224.
141. Goh Y., Tamura T. 1980b. ผลของกรดอะมิโนต่อพฤติกรรมการให้อาหารในลมหายใจทะเลแดง // Sotr. ไบโอชิม. ฟิสิออล. วี 66ซี ป.225-229.
142. Hara T.J., Sveinsson T., Evans R.E., Klaprat D.A. พ.ศ. 2536 ลักษณะทางสัณฐานวิทยาและการทำงานของอวัยวะรับกลิ่นและการรับรสของ Salvelinus สามสายพันธุ์ // Can. เจ ซูล. ว. 71. ลำดับที่. 2. หน้า 414-423.
143. ฮารา ที.เจ. 2537. ความหลากหลายของการกระตุ้นทางเคมีของการดมกลิ่นและความกระวนกระวายใจของปลา // บทวิจารณ์ใน Fish Biol. ปลา. V.4.เลขที่ 1. ป.1-35.
144. ฮาร์ทลีย์ พี.เอช.ที. พ.ศ. 2491 ความสัมพันธ์ด้านอาหารและการให้อาหารในชุมชนปลาน้ำจืด // เจ. แอนิม. อีคอล. ว. 17. ป. 1-14.
145. Hayama T., Caprio J., 1989. โครงสร้าง Lobule และการจัดระเบียบ somatotopic ของกลีบใบหน้าไขกระดูกในช่องปลาดุก Ictalurus punctatus // J. Sotr. นิวรอล. ว.285.เลขที่. 1. ป. 9-17.
146. เฮสเตอร์ เอฟ.เจ. พ.ศ. 2507 ผลของการจัดหาอาหารต่อความดกของไข่ในปลาหางนกยูงตัวเมีย Lebistes reticulatus (Peters) // J. Fish ความละเอียด บด. แคนาดา. ว.21.ลำดับที่. 4. หน้า 757-764.
147. Hidaka I., Yokota S. 1967. การกระตุ้นตัวรับรสชาติด้วยสารรสหวานในปลาคาร์พ // Jpn. เจ. ฟิสิออล. ว. 17. ป. 652-666.
148. Hidaka I., Ohsugi T., Kubomatsu T. 1978. การกระตุ้นตัวรับรสชาติและพฤติกรรมการกินอาหารในปลาปักเป้า Fugu pardalis I. ผลกระทบของสารเคมีชนิดเดียว // ความรู้สึกและรสชาติทางเคมี ว.3.เลขที่. 4. หน้า 341-354.
149. Hidaka I. 1982. การกระตุ้นตัวรับรสชาติและพฤติกรรมการให้อาหารในปลาปักเป้า // ใน: Chemoreception in Fishes (Ed. T. J. Hara) อัมสเตอร์ดัม: Elsevier Scientific Publishing Al. ป.243-257.
150. Hidaka I., Ishida Y. 2528. การตอบสนองอย่างน่ารับประทานใน Shimaisaki (ปลาเสือ) Therapon oxyrhynchus // Bull. ญี่ปุ่น สังคมสงเคราะห์ วิทยาศาสตร์ ปลา. ว. 51. ลำดับที่. 3. หน้า 387-391.
151. Holm J.C., Walther B. 1988. กรดอะมิโนอิสระในแพลงก์ตอนสัตว์น้ำจืดและอาหารแห้ง: ความสำคัญที่เป็นไปได้สำหรับการให้อาหารครั้งแรกในปลาแซลมอนแอตแลนติก fty (Salmo salar) // การเพาะเลี้ยงสัตว์น้ำ ว. 71. ลำดับที่. 3. หน้า 223-234.
152. Horppila J. 1994. อาหารและการเจริญเติบโตของแมลงสาบ (Rutilus rutilus (L.)) ในทะเลสาบ Vesijarvi และการเปลี่ยนแปลงที่เป็นไปได้ในกระบวนการจัดการทางชีวภาพ // Hydrobiologia ว. 294. หน้า 35-41.
153. Horppila J. 1999. การเปลี่ยนแปลงองค์ประกอบอาหารของ Cyprinid ที่กินไม่ได้ทุกชนิดซึ่งเป็นแหล่งที่มาของข้อผิดพลาดที่เป็นไปได้ในการประมาณการบริโภคอาหาร // Hydrobiologia ว. 294. หน้า 35-41.
154. Horppila J., Ruuhijarvi J., Rask M., Karppinen C., Nyberg K., Olin M. 2000. การเปลี่ยนแปลงตามฤดูกาลในอาหารและความอุดมสมบูรณ์ของคอนและแมลงสาบในบริเวณชายฝั่งและทะเลของ ทะเลสาบใหญ่ // เจ.ฟิชไบโอล. ว. 56. ป. 51-72.
155. Humbach I. 1960. Geruch und Geschmack bei den augenlosen HoMenfischen Anoptichthys jordani, Hubbs und Innes und Anoptichtys hubbsi //Naturwissenschaften. บดี 47 ส. 551-560
156. นายพราน เจ.อาร์-.2523 พฤติกรรมการให้อาหารและนิเวศวิทยาของลูกน้ำปลาทะเล//พฤติกรรมของปลาและการใช้ในการจับและเพาะเลี้ยงปลา มะนิลา. ป.287-330.
157. Jakubowski M., Whitear M. 1990. สัณฐานวิทยาเปรียบเทียบและเซลล์วิทยาของต่อมรับรสใน teleosts // Z. mikrosk.-anat. ฟอร์ช. ว.104.เลขที่. 4. หน้า 529-560.
158. โจนส์ เค.เอ. 2532. ความอร่อยของกรดอะมิโนและสารประกอบที่เกี่ยวข้องกับเรนโบว์เทราท์, Salmo gairdneri Richardson // J. Fish Biol. ว.34.เลขที่. 1. หน้า 149-160.
159. โจนส์ เค.เอ. 2533. ข้อกำหนดทางเคมีของการให้อาหารเรนโบว์เทราท์ Oncorhynchus mykiss (Walbaum); การศึกษาความอร่อยของกรดอะมิโน เอไมด์ เอมีน แอลกอฮอล์ อัลดีไฮด์ แซ็กคาไรด์ และสารประกอบอื่นๆ // J. Fish Biol ว.37.ลำดับที่. 3. หน้า 413-423.
160. Kaku T., Tsumagari M., Kiyohara S., Yamashita S. 1980. การตอบสนองอย่างน่ารับประทานในสร้อย Pseudorasbora parva // Physiol. ประพฤติตน ว.25.ลำดับที่. 1. หน้า 99-105.
161. Kanwal J.S. , Caprio J. 1983. การตรวจสอบทางสรีรวิทยาของระบบรับรสของคอหอย (IX-X) ในช่องปลาดุก Ictalurus punctatus // J. Comp.Physiol. ก.V. 150. หน้า 345-357.
162. Kanwal J.S. , Hidaka I. , Caprio J. 1987. การตอบสนองต่อรสชาติต่อกรดอะมิโนจากกิ่งก้านของเส้นประสาทใบหน้าที่ทำให้ต่อมรับรสในช่องปากและนอกช่องปากในช่องปลาดุก Ictalurus punctatus // การวิจัยสมอง ว. 406 ลำดับที่ 1-2. พ. 105112. /
163. Kanwal J.S. , Caprio J. 1988. รสชาติที่ทับซ้อนกันและแผนที่สัมผัสของ oropharynx ในกลีบช่องคลอดของปลาดุกช่องทาง, Ictalurus punctatus // J. Neurobiol ว. ~19. เลขที่ 3. หน้า 211-222.
164. เกษมยาน อ.โอ. พ.ศ. 2538 การตอบสนองการรับกลิ่นและการรับรสของปลาสเตอร์เจียนรุ่นเยาว์และปลาพายต่อสิ่งเร้าทางเคมีตามธรรมชาติและสังเคราะห์ // ใน: Proceeding of International Symp. บนอะซิเพนเซอริดส์ สำนักพิมพ์วีนิโร. มอสโก ป.22-33.
165. Kasumyan A.O. , Sidorov S.S. 1995ก. ผลของน้ำที่เป็นกรดต่อการตอบสนองเชิงพฤติกรรมของเกรย์ลิง ไธมัลลัส ไธมัลลัส ต่อรสชาติสิ่งเร้า // การประชุม SETAC-Europe ครั้งที่ 5, พ.ศ. 2538: วิทยาศาสตร์สิ่งแวดล้อมและระบบนิเวศที่เปราะบาง โคเปนเฮเกน ป.284.
166. Kasumyan A.O. , Sidorov S.S. 1995b. ความอร่อยของกรดอะมิโนอิสระและสารรสชาติคลาสสิกต่อ frolich charr, Salvelinus alpinus erythrinus (Georgi) //Nordic J. Freshwater Res. เลขที่ 71. หน้า 320-323.
167. Kasumyan A.O., Morsy A.M.H., 2541. บทบาทของประสบการณ์การให้อาหารต่อรสนิยมของปลา // Abstr. ระยะที่ 8 อาการ สรีรวิทยาของปลา สวีเดน. อุปซอลา: ม. ป.101.
168. Kawamura G. , Washiyama N. 1989. การเปลี่ยนแปลงทางพันธุกรรมของพฤติกรรมและการสร้างรูปร่างของอวัยวะรับความรู้สึกในเสียงเบสปากใหญ่และปลานิลนิโลติกา // Trans. อาเมอร์. ปลา. โซค.วี. 118. หน้า 203-213.
169. Kiyohara S., Hidaka I., Tamura T. 1975. การตอบสนองอย่างน่ารับประทานในปลาปักเป้า II. วิเคราะห์ไฟเบอร์เดี่ยว//บูล. ญี่ปุ่น สังคมสงเคราะห์ วิทยาศาสตร์ ปลา. ว. 41. ลำดับที่. 4. หน้า 383-391.
170. Kiyohara S., Yamashita S., Harada S. 1981. ความไวสูงของตัวรับรส minnow ต่อกรดอะมิโน // Physiol, และ Behav. ว.26.เลขที่. 6. หน้า 1103-1108. .
171. Kleerekoper, H. 1969. การดมกลิ่นในปลา // สำนักพิมพ์มหาวิทยาลัยอินดนานา. บลูมิงตัน 22 หน้า
172. Klaprat D.A., Evans R.E., Hara T.J. 2535. สารปนเปื้อนในสิ่งแวดล้อมและการรับเคมีบำบัดในปลา // ใน: การรับเคมีบำบัดของปลา (ed. T.J.Hara) แชปนันและฮอลล์ ลอนดอน. ป.321-341.
173. Konishi J., Zotterman Y. 2504. การทำงานของรสชาติในปลาคาร์พ // Acta Physiol. สแกน ว. 52. หน้า 150-161.
174. Konishi J., Zotterman Y. 1963. การทำงานของรสชาติในปลา // ใน: การดมกลิ่นและรสชาติ. อ็อกซ์ฟอร์ด-ปารีส ป.215-233.
175. Konishi J., Niwa H. 1964. คุณสมบัติบางประการของตัวรับรสชาติในปลาน้ำจืด // Jap. เจ. ฟิสิออล. ว. 14. หน้า 328-343.
176. โคนิชิ เจ., ฮิดากะ 1.1969. เรื่องการกระตุ้นตัวรับเคมีของปลาด้วยการเจือจางสารละลายโพลีอิเล็กโตรไลต์ // Jap. เจ. ฟิสิออล. ว. 19. หน้า 315-326.
177. Kumai H., Kimura I., Nakamura M., Takii K., Ishida H. 1989. การศึกษาเกี่ยวกับระบบย่อยอาหารและการดูดซึมของอาหารปรุงแต่งในปลาปักเป้าโอเซลเลต // นิปปอน ซุยซัน กักไกชิ ว. 55. ลำดับที่. 6. หน้า 1,035-1,043.
178. แม็กกี้ A.M. 2525. การจำแนกสารกระตุ้นการกินอาหาร // ใน: Chemoreception ในปลา (Ed. T J. Hara) อัมสเตอร์ดัม: Elsevier Scientific Publishing Al. ป.275-291.
179. Mackie A.M, Mitchell A.I. พ.ศ. 2526 การศึกษาลักษณะทางเคมีของการให้อาหารสารกระตุ้นสำหรับปลาไหลยุโรปรุ่นเยาว์ แองกวิลลา แองกวิลลา (L) // J. Fish Biol V. 22. N. 4. P. 425-430.
180. Marui T., Evans R.E., Zielinski V., Naga T.J. 1983ก. การตอบสนองอย่างน่ารับประทานของเรนโบว์เทราท์ (Salmo gairdneri) ต่อกรดอะมิโนและอนุพันธ์ // J. Sotr. ฟิสิออล. อ.ว. 53.เลขที่ 4. หน้า 423-433.
181. Marui T., Harada S., Kasahara Y. 1983b. ความจำเพาะของรสชาติสำหรับ "กรดอะมิโนในระบบรับรสใบหน้าของปลาคาร์พ, Cyprinus carpio L. // J. Sotr. Physiol. V. 153A. หมายเลข 3. หน้า 299-308
182. Marui T. , Harada S. , Kasahara Y. 2530. กลไกการรับรสที่หลากหลายสำหรับกรดอะมิโนในปลาคาร์พ, Cyprinus carpio L. // Umami: รสชาติพื้นฐาน (eds. Y. Kawamura และ M.R. Kare) นิวยอร์ก: มาร์เซล เดกเกอร์ ป.185199.
183. Marui T., Caprio J. 1992. Teleost gustation // ใน: Fish Chemoreception (ed. T.J.Hara) แชปแมนและฮอลล์ ลอนดอน. ป.171-198.
184. McNicol R.E., Scherer E., Murkin E.J. พ.ศ. 2528 การตรวจสอบภาคสนามเชิงปริมาณของการให้อาหารและพฤติกรรมอาณาเขตของลำธารอายุน้อยแห่งปี, Salvelinus fontinalis // สภาพแวดล้อม ไบโอล ปลา. ว.12. ลำดับที่. 3. หน้า 219229.
185. มัวร์ จี.เอ. พ.ศ. 2493 อวัยวะรับสัมผัสทางผิวหนังของปลาสร้อยมีหนาม ปรับให้เข้ากับชีวิตในน้ำโคลนของภูมิภาค Great Plains // Trans อาเมอร์. มิค. สังคมสงเคราะห์ ว. 69. ป. 69-95.
186. Ohsugi T., Hidaka I., Ikeda M. 1978. การกระตุ้นตัวรับรสชาติและพฤติกรรมการกินอาหารในปลาปักเป้า Fugu pardallis. ครั้งที่สอง ผลที่เกิดจากส่วนผสมของสารสกัดจากหอยลาย // เคม. ความรู้สึกและรสชาติ ว. 3. ลำดับที่. 4. หน้า 355-368.
187. Pausan K. 1984. ตู้ปลา. หนังสือชีวิตชนบท. มิดเดิลเซ็กซ์. 240หน้า
188. Prescott J., Laing D., Bell G., Yoshida M., Gillmore R., Allen S., Yamazaki K., Ishii R. 1992. การตอบสนองแบบ Hedonic ต่อการแก้ปัญหาด้านรสชาติ: การศึกษาข้ามวัฒนธรรมของญี่ปุ่นและออสเตรเลีย // ความรู้สึกทางเคมี ว.17.ลำดับที่. 6. หน้า 801-809.
189. รามิเรซ ไอ. สปรอตต์ อาร์.แอล. พ.ศ. 2521 กลไกทางพันธุกรรมของการดื่มและการให้อาหาร // Neurosci. ไบโอบีฮาฟ สาธุคุณ V.2.เลขที่ 1. ป.15-26.
190. Reid M., Hammersley R. 1996. ผลของเครื่องดื่มอัดลมต่อพฤติกรรมการกินในช่วงเจ็ดวัน // Proc. นูทริ สังคมสงเคราะห์ ว.55.ฉบับที่. 3. หน้า 251.
191. Reutter K. 1971. Die Geschmacksknospen des Zwergwelses Amiurus nebtilosus, Morphologische und histochemixche Unterschungen // Z. mikr. อานนท์. บด. 120. ส. 280-308.
192. Reutter K. 1986. ตัวรับเคมี // ใน: ชีววิทยาของจำนวนเต็ม. V.II. เอ็ด โดย J. Bereiter-Hahn, A. G. Matoltsy และ K. S. Richards) เบอร์ลิน สปริงเกอร์. ป.586-604.
193. Reutter K. 1992. โครงสร้างของอวัยวะรับรส, อัดแน่นโดยปลาไซลูรอยด์ Plotosus lineatus (Thunberg). ใน: Fish Chemoreception (เอ็ด. ใน Chapman and Hall. London. หน้า 60-78.
194. Reutter K., Witt M. 1993. สัณฐานวิทยาของอวัยวะรับรสของสัตว์มีกระดูกสันหลังและการจัดหาเส้นประสาท // ใน: กลไกของการถ่ายโอนรสชาติ (eds. S.A. Simon และ S.D. Roper), CRC Press, Boca Raton ป.29-82.
195. Roper S.D. - 1989. ชีววิทยาของเซลล์ของตัวรับรสของสัตว์มีกระดูกสันหลัง ก. รศ. Neurosci., V. 12, หน้า 329-353.
196. Sakashita H. 1992. พฟิสซึ่มทางเพศและนิสัยการกินของปลา clingfish, Diadesmichthys lineatus และการพึ่งพาอาศัยเม่นทะเลที่เป็นเจ้าภาพ // สภาพแวดล้อม ไบโอล ปลา. ว. 34. ป. 95-101.
197. ซอนเดอร์ส เจ.ที. พ.ศ. 2457 หมายเหตุเกี่ยวกับอาหารปลาน้ำจืด // Proc. .แคมบ. ฟิล. สังคมสงเคราะห์ ว. 17. หน้า 236-239.
198. แซกเซนา เอส.ซี. 2502. อุปกรณ์ติดกาวสำหรับปลาไซปรินิดจากเนินเขา
199. คาร์รา มัลยา // พรอค. แนท. สถาบัน อินเดีย. ว.25.ป.205-214.
200. Saxena S.C., Chandy M. 1966. อุปกรณ์ยึดติดในปลาแม่น้ำจากเนินเขาของอินเดียบางชนิด // J. Zool ว. 148. หน้า 315-340.
201. Schemmel C. 1967. Vergleichende Unterschungen an den Hautsinnesorgane oberund unterirdisch lebender Astyanax-Formen // Z. Morphol. ชั้น. บด. 61. ส. 255-316.
202. Sibbing F.A., Osse J.W.M., Terlouw A. 2529. การจัดการอาหารในปลาคาร์พ (Cyprinus carpio): รูปแบบการเคลื่อนไหว กลไกและข้อ จำกัด // J. Zool สังคมสงเคราะห์ ของลอนดอน ว. 210(ก) เลขที่ 2. หน้า 161-203.
203. ซินแคลร์ เจ.ดี., คัมปอฟ-โพเลวอย เอ., สจ๊วร์ต อาร์., ลี ที.-เค. พ.ศ. 2535 รสนิยมในบรรทัดหนูที่เลือกสำหรับการบริโภคแอลกอฮอล์ต่ำและสูง // แอลกอฮอล์ ว.9.เลขที่. 2. หน้า 155-160.
204. ชิงไก ต., ไบด์เลอร์ แอล.เอ็ม. พ.ศ. 2528 ยับยั้งความแตกต่างในการตอบสนองต่อรสขมในหนู // ความรู้สึกทางเคมี ว.10.ลำดับที่. 1. น.51-55.
205. Specziar A., Tolg L., Biro P. 1997. กลยุทธ์การให้อาหารและการเจริญเติบโตของไซปรินิดส์ในเขตชายฝั่งของทะเลสาบบาลาตัน // J. Fish Biol. ว. 51. หน้า 11091124.
206. Stradmeyer L., Metcalfe N.B., Thorpe J.E. 2531. ผลของรูปร่างเม็ดอาหารและเนื้อสัมผัสต่อการตอบสนองการให้อาหารของปลาแซลมอนแอตแลนติกรุ่นเยาว์ // การเพาะเลี้ยงสัตว์น้ำ. V. 73. หน้า 217-228.
207. Takeda M., Takii K., Matsui K. 1984. การจำแนกสารกระตุ้นการให้อาหารสำหรับปลาไหลเด็กและเยาวชน // บูล. ญี่ปุ่น สังคมสงเคราะห์ วิทยาศาสตร์ ปลา. ว. 50. ลำดับที่. 4. ป. 645651 ป
208. Takeda M., Takii K. 1992. รสชาติและโภชนาการในปลา: การประยุกต์กับการเพาะเลี้ยงสัตว์น้ำ // ใน: การรับเคมีบำบัดของปลา (Ed. T.J.Hara) แชปแมนและฮอลล์ ลอนดอน. ป.271-287.
209. Thorman S. 1982. พลวัตเฉพาะและการแบ่งทรัพยากรในสมาคมปลาที่อาศัยอยู่บริเวณปากแม่น้ำตื้นบนชายฝั่งตะวันตกของสวีเดน // Oikos ว.39.ป.32-39.
210. ธอร์มัน เอส., วีเดอร์โฮล์ม อ.เอ็ม. 2526. การเกิดตามฤดูกาลและการใช้อาหารของการรวมตัวของพันธุ์ปลาชายฝั่งทะเลในทะเลบอทเนีย. สวีเดน // มี.ค. เอคอล โครงการ เซอร์ ว. 10. ป. 223-229.
211. ทริเวอร์ส อาร์.แอล. 2515. การลงทุนของผู้ปกครองและการเลือกเพศ. ใน: การเลือกเพศและการสืบเชื้อสายมาของมนุษย์ (บรรณาธิการ บี.จี. แคมป์เบลล์) อัลดีน. ชิคาโก. ป.136-179.
212. เวเธอร์ลีย์ เอ็น.เอส. พ.ศ. 2530 การรับประทานอาหารและการเจริญเติบโตของ dace 0 กลุ่ม Leuciscus leucis"cus (L. ) และแมลงสาบ Rutilus rutilus (L. ) ในแม่น้ำที่ราบลุ่ม // J. Fish Biol V. 30. P. 237- 247.
213. Weinrich M.T., Schilling M.R., Belt C.R. พ.ศ. 2535 หลักฐานสำหรับการได้มาซึ่งพฤติกรรมการกินอาหารแบบใหม่: (ต้องให้อาหารวาฬหลังค่อม, Megaptera novaengliae) // Anim ประพฤติตน ว. 44. ลำดับที่. 6. หน้า 1,059-1,072.
214. Witt M., Reutter K. 1990. การสาธิตด้วยกล้องจุลทรรศน์อิเล็กตรอนของบริเวณจับกับเลคตินในต่อมรับรสของปลาดุกยุโรป Silurus glanis (Teleostei) // ฮิสโตเคมี. ว. 94. หน้า 617-628.
215. White A.W., Fukuhara O., Anraku M.1989. อัตราการตายของปลา lan/ae จากการกินสารพิษไดโนแฟลเจลเลตที่เป็นพิษ // กระแสน้ำแดง: ชีววิทยา สิ่งแวดล้อม 116 วิทยาศาสตร์ และพิษวิทยา โปรค อินเตอร์ที่ 1 อาการ กระแสน้ำสีแดง นิวยอร์ก. ป.395398!
216. วูตตัน อาร์.เจ. พ.ศ. 2519 ชีววิทยาของ Sticklebacks // สำนักพิมพ์วิชาการ. ลอนดอน. นิวยอร์ก. ซานฟรานซิสโก. ภาควิชาสัตววิทยา. เวลส์ ป.57371.
217. Yamaguchi S. 1991. คุณสมบัติพื้นฐานของอูมามิและผลกระทบต่อมนุษย์ // Physiol. ประพฤติตน ว. 49. หน้า 833-841.
218. Yoshii K., Kato N., Kurihara K., Kabatake Y. 1979. การตอบสนองอย่างลิ้นจี่ของตัวรับเพดานปากปลาไหลต่อกรดอะมิโนและกรดคาร์บอกซิลิก // J. Gen. ฟิสิออล. ว. 74. ลำดับที่. 3. หน้า 301-317.
ระบบรสชาติใช้ในการประเมิน รสชาติของผลิตภัณฑ์– ความรู้สึกที่ซับซ้อนที่รับรู้เมื่อถูกทดสอบ – ชิม (การประเมินผลิตภัณฑ์อาหารในช่องปาก)
รสชาติ – ความรู้สึกที่เกิดขึ้นเมื่อกระตุ้นต่อมรับรสและถูกกำหนดทั้งเชิงคุณภาพและเชิงปริมาณ
การประเมินรสชาติขึ้นอยู่กับการกำหนดประเภทของรสชาติและความเข้มข้นของรสชาติ มาตรฐานของสารรับรสปฐมภูมิในวิทยาศาสตร์สินค้าโภคภัณฑ์ถือเป็นความหวานตามลำดับ – ซูโครส; เปรี้ยว – กรดไฮโดรคลอริก; เค็ม – เกลือ; ขม – ควินิน (คาเฟอีน) สามารถรับรสชาติประเภทและเฉดสีอื่นๆ ทั้งหมดได้โดยการผสมรสชาติหลักที่เป็นไปได้สามในสี่ตามสัดส่วนที่ต้องการ
การกำหนดรสชาติเชิงคุณภาพนั้นเกิดจากผลของสารที่มีต่อต่อมรับรสซึ่งอยู่ที่ลิ้นเป็นหลัก นอกจากนี้ยังพบบนพื้นผิวเมือกของช่องปาก ผนังคอหอย ต่อมทอนซิล และกล่องเสียง จำนวนต่อมรับรสในช่องปากของมนุษย์มีจำนวนถึง 9,000 นอกจากนี้การกำหนดรสชาติยังสัมพันธ์กับการสัมผัสอาหารในช่องปากอีกด้วย
อุปกรณ์รับรสในปากของมนุษย์เป็นเครื่องวิเคราะห์ทางเคมีและมีความไวมากกว่าเครื่องมือสมัยใหม่ ความสมบูรณ์ของเฉดสีต่างๆ และการผสมผสานของความรู้สึกรับรสเกิดขึ้นอันเป็นผลมาจากการระคายเคืองของอวัยวะสัมผัสพิเศษ – ปุ่มรับรส (ปุ่ม) ซึ่งแต่ละปุ่มประกอบด้วยเซลล์รับเคมีที่ไวต่อความรู้สึกหลายเซลล์ที่เชื่อมต่อกับเซลล์ประสาทรับความรู้สึก เซลล์รับเคมีตอบสนองต่อสารเคมีบางชนิด
ปุ่มรับรสมีความแตกต่างกันเพื่อรับรู้ถึงรสชาติหลักๆ ได้แก่ หวาน เค็ม เปรี้ยว และขม หัวที่ปลายลิ้นไวต่อรสหวานที่สุดที่ขอบด้านหน้าของลิ้น - ถึงรสเค็มที่ขอบด้านหลังลิ้น – เปรี้ยวที่ฐาน – ขมขื่น
สารทั้งหมดที่กำหนดรสชาติของอาหารจะละลายได้ในน้ำ เฉพาะในรูปแบบที่ละลายเท่านั้นที่จะส่งผลต่อเครื่องวิเคราะห์ทางเคมีของเครื่องรับรส
เกณฑ์การรับความรู้สึกยังขึ้นอยู่กับอุณหภูมิของสารละลาย ซึ่งอาจอธิบายได้จากการเปลี่ยนแปลงสถานะของโมเลกุลโปรตีนปุ่มรับรส การรับรู้สารปรุงแต่งรสที่ดีที่สุดเกิดขึ้นที่อุณหภูมิสารละลายใกล้กับอุณหภูมิร่างกายมนุษย์ (36.5 ° C) สารละลายร้อนของสารเดียวกันในระดับความเข้มข้นที่ระบุดูไม่มีรสนั่นคือไม่ทำให้เกิดความรู้สึกใด ๆ เมื่อเย็นลงถึง 30°C รสหวานจะพัฒนามากกว่ารสเค็มหรือรสขม
เมื่อประเมินรสชาติ ความเร็วของการรับรู้รสชาติก็มีความสำคัญเช่นกัน รสเค็มรับรู้ได้เร็วที่สุด รสหวานและเปรี้ยวรับรู้ได้ช้ากว่า รสขมจะรับรู้ได้ช้าที่สุด
ลักษณะรสชาติดังต่อไปนี้มีความโดดเด่น
รสเปรี้ยว--ระบุลักษณะรสชาติพื้นฐานของสารละลายน้ำของกรดส่วนใหญ่ (เช่นกรดซิตริกและทาร์ทาริก) ความรู้สึกรับกลิ่นและการรับรสที่ซับซ้อนซึ่งมีสาเหตุหลักมาจากการมีกรดอินทรีย์
ความเปรี้ยว – คุณสมบัติทางประสาทสัมผัสของสารหรือสารผสมแต่ละชนิดที่ทำให้เกิดรสเปรี้ยว
รสเปรี้ยว-- แสดงถึงระดับความเข้มข้นของรสชาติที่ต่ำกว่าของผลิตภัณฑ์รสเปรี้ยว
รสขม--แสดงถึงลักษณะรสชาติพื้นฐานที่เกิดจากสารละลายในน้ำของสารเคมี เช่น ควินินและคาเฟอีน รวมถึงอัลคาลอยด์บางชนิด
ความขมขื่น – คุณสมบัติทางประสาทสัมผัสของสารหรือสารผสมแต่ละชนิดที่ทำให้เกิดรสขม
รสเค็ม--แสดงถึงลักษณะรสชาติพื้นฐานที่เกิดจากสารละลายน้ำของสารเคมี เช่น โซเดียมคลอไรด์
ความเค็ม – คุณสมบัติทางประสาทสัมผัสของสารเดี่ยวหรือสารผสมที่ทำให้เกิดรสเค็ม
รสหวาน --แสดงถึงลักษณะรสชาติพื้นฐานที่เกิดจากสารละลายในน้ำของสารเคมี เช่น ซูโครส
ความหวาน – คุณสมบัติทางประสาทสัมผัสของสารเดี่ยวหรือสารผสมที่ทำให้เกิดรสหวาน
รสชาติอัลคาไลน์--แสดงลักษณะรสชาติพื้นฐานที่เกิดจากสารละลายน้ำของสารเคมี เช่น โซเดียมไบคาร์บอเนต
ความเป็นด่าง – คุณสมบัติทางประสาทสัมผัสของสารเดี่ยวหรือสารผสมที่ทำให้เกิดรสชาติเป็นด่าง
รสฝาดรสเปรี้ยว -แสดงถึงความรู้สึกที่ซับซ้อนที่เกิดจากการหดตัวของพื้นผิวเมือกของช่องปากและเกิดขึ้นจากอิทธิพลของสารเช่นแทนนิน ความฝาด – คุณสมบัติทางประสาทสัมผัสของสารหรือสารผสมแต่ละชนิดที่ทำให้เกิดรสฝาด
รสชาติโลหะ--แสดงถึงลักษณะรสชาติพื้นฐานที่เกิดจากสารละลายน้ำของสารเคมี เช่น เฟอร์รัสซัลเฟต
รสจืด จืดชืด – ผลิตภัณฑ์ที่ไม่มีรสชาติเฉพาะตัว
รสที่ค้างอยู่ในคอ – ความรู้สึกรับรสที่ปรากฏหลังจากกลืนหรือเอาผลิตภัณฑ์ออกจากช่องปาก ซึ่งแตกต่างจากความรู้สึกที่รับรู้ขณะอยู่ในช่องปาก
ผลิตภัณฑ์อาหารมีรสชาติเดียว (น้ำตาล – หวานเกลือแกง – เค็มกรด – เปรี้ยว) หรือแตกต่างกันในรสชาติประเภทพื้นฐานรวมกัน ในกรณีนี้เราสามารถพูดคุยเกี่ยวกับการผสมผสานรสนิยมที่กลมกลืนและไม่ลงรอยกัน . ใช่แล้ว ก อย่างกลมกลืนโดยรวมแล้วมีรสหวานหรือรสเค็มผสมกับรสเปรี้ยวหรือขม เช่น รสหวานอมเปรี้ยวของผลไม้ ขนมหวาน รสหวานปนขมของช็อกโกแลต ผักดองรสเค็ม-เค็ม มะกอกรสเค็ม-ขม ไม่ลงรอยกันพิจารณาส่วนผสมต่างๆ เช่น เค็ม-หวาน ขม-เปรี้ยว การรวมกันเหล่านี้ถูกมองว่าเป็นสองรสชาติที่แตกต่างกันซึ่งเป็นเรื่องปกติสำหรับผลิตภัณฑ์อาหารหายากและเกิดขึ้นตามกฎซึ่งเป็นผลมาจากการเน่าเสีย (เช่นรสเปรี้ยวอมขมของผักดอง)
การรับรู้รสชาติขึ้นอยู่กับองค์ประกอบทางเคมี ความหนืด และปริมาณของอาหาร ลักษณะของสารที่มีกลิ่นและรสชาติและความเข้มข้นของการปล่อย, อัตราการกำจัด, ลักษณะของผลกระทบต่ออวัยวะของการรับรส; ความพร้อมใช้งานของสารเหล่านี้ในช่วงเวลาหนึ่ง เงื่อนไขในการรับประทานอาหาร (โดยเฉพาะการหายใจ ปริมาณและความเร็วของการไหลของน้ำลาย ระยะเวลา อุณหภูมิ) และคุณภาพการเคี้ยวผลิตภัณฑ์
เป็นที่ยอมรับว่าการประเมินความเข้มข้นของรสชาติพื้นฐานอาจได้รับอิทธิพลจากสีของผลิตภัณฑ์ ดังนั้นสีเหลืองและสีเขียวอ่อนจะเพิ่มการประเมินความเข้มของความเป็นกรดของผลิตภัณฑ์ และสีแดงจะช่วยเพิ่มการประเมินความเข้มของความหวานเมื่อเทียบกับสีที่ไม่มีสี คุณควรคำนึงถึงช่วงเวลาที่รสชาติหนึ่งถูกระงับโดยอีกรสชาติหนึ่งด้วย เช่น รสเปรี้ยวถูกระงับด้วยรสหวาน และในระดับที่น้อยกว่า – เค็มและขม รสเค็มและขมถูกระงับโดยซูโครสและกรดซิตริกที่ความเข้มข้นบางอย่าง รสหวานถูกระงับเล็กน้อยด้วยกรดซิตริกที่มีความเข้มข้นเล็กน้อย
รสที่คงอยู่แม้ความอยากอันเป็นเหตุให้ดับไปนั้นเรียกว่า รสชาติรอง . มันอาจจะเหมือนกันหรือตรงกันข้ามก็ได้ เดียวกันมันเกิดขึ้นเพราะว่าหลังจากการสิ้นสุดของแรงกระตุ้นแห่งรสชาติแล้ว ความรู้สึกในรสชาติก็ยังคงมีคุณภาพเหมือนเดิมกับความรู้สึกที่มีอยู่ในระหว่างอิทธิพลของแรงกระตุ้นของรสชาติ ตัดกันเรียกว่ารสทุติยภูมิซึ่งเกิดขึ้นหลังจากการขจัดแรงกระตุ้นรสชาติที่ใช้งานอยู่
ผลิตภัณฑ์อาหารทุกชนิดทำให้เกิดรสชาติรองบางอย่าง ไม่ว่าจะเหมือนกันหรือต่างกันก็ตาม หากรสชาติรองมีความสม่ำเสมอและตรงกับโปรไฟล์รสชาติหลักของผลิตภัณฑ์และหายไปอย่างรวดเร็วเมื่อกลืนชิ้นส่วนนั้นลงไป นั่นพิสูจน์ได้ว่าคุณภาพของผลิตภัณฑ์อยู่ในระดับสูง แต่หากหลังจากกลืนแล้วยังมีรสชาติรองอยู่ในปากแสดงว่าผลิตภัณฑ์นั้นด้อยกว่ามูลค่าผู้บริโภคเป็นอันดับแรก ในการฝึกวิเคราะห์ทางประสาทสัมผัสมักพบรสชาติที่ตัดกันรองเช่นน้ำกลั่นหลังจากบ้วนปากด้วยสารละลายเกลือแกงดูเหมือนมีรสหวาน หลังจากชิมรสหวานได้ชั่วครู่ รสเปรี้ยวก็รับรู้ได้ชัดเจนยิ่งขึ้น และความรู้สึกไม่พึงประสงค์ก็รุนแรงขึ้น ดังนั้นไวน์บ่มจึงไม่ได้รับการจัดอันดับตามไวน์หวาน อย่าประเมินผลิตภัณฑ์ที่มีเกลือเล็กน้อยหลังจากประเมินผลิตภัณฑ์ที่มีเกลือแห้ง
ความอร่อย(รส– รสชาติ) – ความรู้สึกที่ซับซ้อนในปากที่เกิดจากรสชาติ กลิ่น และเนื้อสัมผัสของผลิตภัณฑ์อาหาร โดยพิจารณา (ประเมิน) ทั้งเชิงคุณภาพและเชิงปริมาณ
การรับรสอาจได้รับอิทธิพลจากการสัมผัส ความร้อน ความเจ็บปวด และ/หรือความรู้สึกทางการเคลื่อนไหวร่างกาย
การประเมินคุณภาพผลิตภัณฑ์อาหารโดยอาศัยการรับรู้ที่ซับซ้อนและการวิเคราะห์ทางการมองเห็น การดมกลิ่น สัมผัส การรับรส เสียง และแรงกระตุ้นอื่นๆ (สิ่งกระตุ้น) เรียกว่าการประเมินความละเอียดอ่อนหรือรสชาติ เมื่อเทียบกับรสชาติของมัน
การเปลี่ยนแปลงเชิงปริมาณในองค์ประกอบแต่ละส่วนของความอร่อยในช่วงเวลาหนึ่งนำไปสู่การก้าวกระโดดเชิงคุณภาพและผลลัพธ์ที่ได้คือผลิตภัณฑ์คุณภาพสูงที่มีรสชาติที่กลมกลืนหรือสมบูรณ์ เมื่อเวลาผ่านไป ความสมดุลระหว่างส่วนประกอบแต่ละส่วนของความอร่อยจะถูกทำลายลง ส่งผลให้คุณภาพของผลิตภัณฑ์ลดลง ตัวอย่างคือกระบวนการสุก การแก่ และการตายของไวน์
เมื่อประเมินรสชาติ จำเป็นต้องคำนึงถึงปรากฏการณ์ที่เกิดจากลักษณะทางสรีรวิทยาของอวัยวะรับสัมผัส เช่น การปรับตัวและความเหนื่อยล้า
การปรับตัว – นี่คือการลดลงของความไวของอวัยวะรับรสซึ่งเกิดจากการสัมผัสกับแรงกระตุ้นรสชาติที่มีคุณภาพเท่ากันและความเข้มข้นคงที่เป็นเวลานาน เมื่อทำการทดสอบตัวอย่างจำนวนมากที่มีรสชาติและความเข้มข้นเท่ากัน การปรับเปลี่ยนจะทำให้เกิดผลลัพธ์ที่บิดเบี้ยว อวัยวะรับรสซึ่งตรงกันข้ามกับการมองเห็นและคล้ายกับการรับรู้กลิ่นนั้นมีลักษณะเฉพาะคือการปรับตัวที่รวดเร็ว
ความเหนื่อยล้า– ลดการรับรู้รสชาติเนื่องจากความเมื่อยล้าของต่อมรับรสภายใต้อิทธิพลของแรงกระตุ้นซ้ำ ๆ โดยจะเกิดขึ้นหลังจากเวลาที่แตกต่างกัน ขึ้นอยู่กับคุณสมบัติของผลิตภัณฑ์ สภาพทางสรีรวิทยาและจิตวิทยาของผู้ทดสอบ การฝึกอบรม และสภาพการทำงาน
480 ถู | 150 UAH | $7.5 ", เมาส์ออฟ, FGCOLOR, "#FFFFCC",BGCOLOR, #393939");" onMouseOut="return nd();"> วิทยานิพนธ์ - 480 RUR จัดส่ง 10 นาทีตลอดเวลา เจ็ดวันต่อสัปดาห์และวันหยุด
อิซาวา โอลกา มิคาอิลอฟนา ความชอบด้านรสชาติและพฤติกรรมการรับรสของปลาคาร์พ: วิทยานิพนธ์... ผู้สมัครสาขาวิทยาศาสตร์ชีวภาพ: 03.00.10.- มอสโก, 2550.- 171 น.: ป่วย อาร์เอสแอล โอดี, 61 07-3/1170
การแนะนำ
บทที่ 1 การทบทวนวรรณกรรม 10
บทที่ 2 วัสดุและวิธีการ 3 8
2.1. วัตถุประสงค์การวิจัย 3 8
2.2. เงื่อนไขการเก็บรักษาและการเตรียมปลาสำหรับการทดลอง 39
2.3. การกำหนดความชอบด้านรสชาติ 40
2.4. การศึกษาพลวัตของการตอบสนองรสนิยมทางพฤติกรรม 42
2.5. การเตรียมแกรนูลทดลอง 43
2.6. ลักษณะทั่วไปของวัสดุ 46
บทที่ 3 การตอบสนองพฤติกรรมการกินของปลาไซปรินิดต่อสารปรุงแต่งรสดั้งเดิม กรดอะมิโนอิสระ และกรดอินทรีย์ 48
3.1. ลิน 48
3.1.1. รสคลาสสิก 48
3.1.2. กรดอะมิโนอิสระ 49
3.1.3. กรดอินทรีย์ 5 0
3.2. กอร์ชัก 55
3.2.1. รสคลาสสิก 5 5
3.2.2. กรดอะมิโนอิสระ 55
3.2.3. กรดอินทรีย์ 57
3.3. เวอร์คอฟกา 60
3.3.1. รสคลาสสิค 60
3.3.2. กรดอะมิโนอิสระ 60
3.4. ปลาคาร์ปทอง64
3.4.1. รสคลาสสิค 64
3.4.2. กรดอะมิโนอิสระ 65
3.5.1. รสคลาสสิค 68
3.5.2. กรดอะมิโนอิสระ 69
บทที่ 4 พลวัตของการสำแดงการตอบสนองพฤติกรรมการรับรสของปลาไซปรินิด 72
4.1. ลิน 72
4.2.1. ซีสเตอีน 76 เม็ด
4.2.2. กลูตามีน 81 เม็ด
4.3. กอร์ชัก 84
บทที่ 5 การอภิปรายผล 86
บทสรุป 123
ข้อสรุป 127
รายชื่อวรรณกรรมที่อ้างถึง
ความรู้เบื้องต้นเกี่ยวกับงาน
ความเกี่ยวข้องของหัวข้อ. พฤติกรรมการให้อาหารเป็นรากฐานของหน้าที่ที่สำคัญที่สุดของอาหารปลา ซึ่งก็คือโภชนาการ ซึ่งมีความสำคัญอย่างยิ่งต่อทั้งต่อบุคคลและประชากรและสายพันธุ์โดยรวม บทบาทนำในการสนับสนุนทางประสาทสัมผัสในช่วงสุดท้ายของพฤติกรรมการให้อาหารปลานั้นมีบทบาทโดยการรับรสชาติ ซึ่งรับประกันการประเมินคุณสมบัติด้านรสชาติของเหยื่อและสอดคล้องกับความต้องการทางโภชนาการของปลาตลอดจนการบริโภคอาหารที่เพียงพอ รายการ (Atema, 1980; Pavlov, Kasumyan, 1990, 1998; Kasumyan, 1997 ) จนกระทั่งเมื่อไม่นานมานี้ การศึกษาจำนวนมากเกี่ยวกับระบบรสชาติของปลามุ่งเน้นไปที่การอธิบายสัณฐานวิทยาและภูมิประเทศของปุ่มรับรสเป็นหลัก ศึกษาโครงสร้างพิเศษและการปกคลุมด้วยเส้น และศึกษาการจัดโครงสร้างทางสัณฐานวิทยาของส่วนกลางของระบบรับรส (Zuwala, Jakubowski, 1993; รอยเตอร์, 1992; Jakubowski, Zuwala, 2000 ; Vasilevskaya, 1974) การศึกษาจำนวนมากเกี่ยวกับคุณสมบัติเชิงหน้าที่ของระบบรสชาติของปลาดำเนินการโดยใช้วิธีอิเล็กโทรสรีรวิทยาเป็นหลัก (Mati และ Caprio, 1992; Jones, 1990; Sutterlin, 1975) อย่างไรก็ตามเมื่อเร็ว ๆ นี้ทิศทางอื่นในการศึกษาการรับรู้รสชาติได้รับการพัฒนาอย่างรวดเร็วและมีประสิทธิผลมาก: นี่เป็นวิธีทดสอบปฏิกิริยาเชิงพฤติกรรมด้วยความช่วยเหลือซึ่งคุณสามารถประเมินความชอบด้านรสชาติของปลาจำนวนมากได้ ปริมาณข้อมูลการทดลองที่สะสมในช่วงไม่กี่ปีที่ผ่านมาทำให้มีแนวคิดที่ชัดเจนเกี่ยวกับรูปแบบทั่วไปและคุณลักษณะเฉพาะของความสัมพันธ์ระหว่างปลากับสิ่งเร้าในรสชาติ ความเหมือนและความแตกต่างในสเปกตรัมอาหารของสารปรุงแต่งรสที่มีประสิทธิภาพในปลาทุกวัยและเป็นระบบ ตำแหน่ง และยังแสดงความเหมือนและความแตกต่างระหว่างการรับรสภายนอกและในช่องปากด้วย (Kasumyan, 1997; Kasumyan, D/aving, 2003)
แต่จนถึงขณะนี้คำถามของการมีอยู่ของความคล้ายคลึงและ/หรือความแตกต่างในการตั้งค่ารสนิยมของความสัมพันธ์อย่างใกล้ชิด
ชนิดของปลา ไม่มีข้อมูลเกี่ยวกับลักษณะทางสรีรวิทยาของการแสดงการตอบสนองต่อพฤติกรรมการรับรสในปลา ไม่มีข้อมูลเกี่ยวกับอิทธิพลของวิถีชีวิตของปลาไซปรินิดสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิดต่อการแสดงออกขององค์ประกอบต่าง ๆ ของการตอบสนองเชิงพฤติกรรมต่อสิ่งเร้ารสชาติต่างๆ
การศึกษาประเด็นเหล่านี้และประเด็นสำคัญอื่นๆ เกี่ยวกับการรับรู้รสชาติของปลามีความสำคัญทางทฤษฎี นอกเหนือจากการปฏิบัติจริงแล้ว ทำให้สามารถชี้แจงบทบาทของระบบประสาทสัมผัสนี้ในการรับรองการเลือกให้อาหารปลาด้วยรายการอาหารที่เพียงพอและในการรักษาสภาวะสมดุล ความรู้เกี่ยวกับรูปแบบความไวต่อรสชาติของปลา ลักษณะการตอบสนองต่อสารปรุงแต่งชนิดต่างๆ มีความสัมพันธ์กับความเป็นไปได้ในการแก้ไขปัญหาเร่งด่วนของการเพาะเลี้ยงสัตว์น้ำสมัยใหม่ เช่น การค้นหาสารกระตุ้นทางโภชนาการทางเคมีที่มีประสิทธิภาพสูง การปรับปรุงที่มีอยู่และการสร้าง อาหารเทียมชนิดใหม่ การพัฒนาเทคโนโลยีชีวภาพสำหรับการเลี้ยงปลา ตลอดจนเหยื่อเคมีเทียมและเหยื่อปลา
เป้าหมายของการทำงาน เพื่อศึกษาความชอบด้านรสชาติ ความไวต่อสิ่งเร้ารสชาติประเภทต่างๆ และลักษณะของพฤติกรรมการรับรสในปลาสายพันธุ์ที่เกี่ยวข้องกันอย่างใกล้ชิด (โดยใช้ตัวอย่างปลาในตระกูลปลาคาร์พ Cyprinidae)
วัตถุประสงค์ของการศึกษาประกอบด้วย:
เปรียบเทียบความชอบด้านรสชาติของสารปรุงแต่งรสดั้งเดิมกับกรดอะมิโนอิสระในปลาไซปรินิด
ค้นหารสชาติของกรดอินทรีย์ในปลาคาร์พ
สำรวจความสัมพันธ์ระหว่างรสชาติที่น่าดึงดูดของสารกับคุณสมบัติทางโครงสร้างและเคมีกายภาพบางประการ
กำหนดระดับความไวต่อรสชาติของปลาคาร์พต่อสารปรุงแต่งที่มีคุณสมบัติกระตุ้นและยับยั้ง
เพื่อศึกษาพฤติกรรมการกินของปลาคาร์พ โครงสร้าง พลวัต และลักษณะเฉพาะขององค์ประกอบหลักของการตอบสนองพฤติกรรมการกินของปลาคาร์พ
ความแปลกใหม่ทางวิทยาศาสตร์ ความแปลกใหม่ทางวิทยาศาสตร์ของงานวิทยานิพนธ์นี้อยู่ที่การขยายความเข้าใจลักษณะการทำงานของระบบรสชาติในปลาสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิด และอธิบายลักษณะของการตอบสนองพฤติกรรมรสชาติที่พวกมันแสดงออกมา งานนี้เป็นครั้งแรกที่กำหนดรสนิยมของปลาคาร์พ 5 สายพันธุ์สำหรับสารปรุงแต่งรสคลาสสิกและกรดอะมิโนอิสระ และปลา 2 สายพันธุ์สำหรับกรดอินทรีย์
เป็นครั้งแรกที่มีการกำหนดระดับความไวต่อรสชาติของปลาคาร์พ 3 สายพันธุ์ต่อสารที่มีคุณสมบัติด้านรสชาติเชิงบวก รวมถึงสารที่ทำให้เกิดการตอบสนองต่อรสชาติเชิงลบ (สารยับยั้ง) แสดงให้เห็นว่าไม่มีความเชื่อมโยงที่เหมือนกันกับปลาคาร์พระหว่างระดับความน่าดึงดูดของรสชาติของสารและคุณสมบัติทางเคมีกายภาพ (น้ำหนักโมเลกุล ค่า pH ของสารละลาย จำนวนกลุ่มฟังก์ชัน) เป็นครั้งแรกที่มีการศึกษาโครงสร้างของการตอบสนองรสชาติเชิงพฤติกรรมของปลาโดยใช้โปรแกรมคอมพิวเตอร์ดั้งเดิม "VN-Fish" พลวัตของการแสดงออกได้รับการชี้แจงและระยะเวลาของการกระทำตามพฤติกรรมแต่ละอย่างของการตอบสนองต่อรสชาติถูกกำหนดแยกต่างหากสำหรับ การทดลองที่สิ้นสุดด้วยการกินหรือการปฏิเสธวัตถุอาหารเทียม (อาหารเม็ด)
ความสำคัญเชิงปฏิบัติของงาน ผลการศึกษานี้สามารถนำไปใช้ในการปฏิบัติงานด้านการเพาะเลี้ยงสัตว์น้ำและการประมงในการค้นหาสารกระตุ้นเคมีอาหารที่มีประสิทธิภาพสูง ในการปรับปรุงและพัฒนาเหยื่อและเหยื่อเทียมในการเตรียมและปรับปรุงสูตรอาหาร
อาหารเทียมเพื่อเพิ่มความอร่อยให้กับปลา ผลลัพธ์ที่ได้รับจากพลวัตของการตอบสนองพฤติกรรมรสชาติทำให้เรามีความคิดที่ชัดเจนยิ่งขึ้นเกี่ยวกับลักษณะของการแสดงออกของพฤติกรรมการให้อาหารที่จัดอย่างซับซ้อนโดยปลาในระยะสุดท้ายและวิธีการควบคุมโดยตรงโดยใช้สิ่งกระตุ้นทางเคมี ผลการศึกษานี้ใช้เป็นส่วนหนึ่งของหลักสูตรการบรรยาย "สรีรวิทยาของปลา" ซึ่งมอบให้กับนักศึกษาภาควิชา Ichthyology คณะชีววิทยา มหาวิทยาลัยแห่งรัฐมอสโก บทบัญญัติที่ได้รับการคุ้มครอง
ความชอบด้านรสชาติของปลาสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิดซึ่งอยู่ในตระกูลเดียวกัน (ไซปรินิด) มีคุณลักษณะเฉพาะจากความจำเพาะของสายพันธุ์สูง
ความอร่อยของสารบางชนิดอาจจะเหมือนกันในปลาสายพันธุ์ที่เกี่ยวข้องกันอย่างใกล้ชิด
ความเข้มข้นของสารที่มีคุณสมบัติน่าดึงดูดและน่ารังเกียจสำหรับปลานั้นใกล้เคียงกัน
พฤติกรรมการกินของปลามีโครงสร้างและไดนามิกที่แน่นอน โดยมีลักษณะเฉพาะและเหมือนกันในปลาที่มีวิถีชีวิตและพฤติกรรมการกินอาหารที่แตกต่างกัน และขึ้นอยู่กับคุณสมบัติด้านรสชาติของวัตถุที่เป็นอาหาร
งานเสร็จแล้ว ด้วยการสนับสนุนทางการเงินจากมูลนิธิรัสเซียเพื่อการวิจัยขั้นพื้นฐาน (ทุน 04-04-48157 และ 07-04-00793) และอยู่ภายใต้กรอบแผนงานเฉพาะเรื่องของสถาบันวิทยาศาสตร์แห่งรัฐ "NIIERV"
การอนุมัติงาน วิทยานิพนธ์ถูกนำเสนอในการประชุม All-Russian Symposium "อายุและสรีรวิทยานิเวศวิทยาของปลา" (Borok, IBVV, 1998), การประชุมระหว่างมหาวิทยาลัยนานาชาติ "Lomonosov-98" (มอสโก, มหาวิทยาลัยแห่งรัฐมอสโก, 1998), การประชุมระหว่างมหาวิทยาลัยครั้งที่ 2 อุทิศให้กับพื้นที่ชุ่มน้ำวันอนุรักษ์น้ำโลก (Rybnoe, 1999), การประชุมจริยธรรมระหว่างประเทศครั้งที่ 26 (บังกาลอร์, อินเดีย, 1999), การประชุมนานาชาติ “การเชื่อมต่อทางโภชนาการในน้ำ
ชุมชนและระบบนิเวศ" (Borok, 2003), การประชุมทางวิทยาศาสตร์ระดับนานาชาติครั้งที่สอง "เทคโนโลยีชีวภาพ - การคุ้มครองสิ่งแวดล้อม" (มอสโก, 2004), การประชุมระดับนานาชาติ "ปัญหาสมัยใหม่ของสรีรวิทยาและชีวเคมีของสิ่งมีชีวิตทางน้ำ" (Petrozavodsk, 2004), การประชุมระดับนานาชาติ "พฤติกรรมปลา " (Borok, 2005), สภาหอดูดาวแห่งรัฐครั้งที่ 9 ของ Russian Academy of Sciences (Tolyatti, 2006), การประชุมนานาชาติ "ปัญหานิเวศวิทยาประชากรของสัตว์" (Tomsk, 2006), การประชุมนานาชาติ IV "การสื่อสารทางเคมีของสัตว์" ปัญหาพื้นฐาน" (มอสโก, 2549) ที่การประชุมเชิงปฏิบัติการของห้องปฏิบัติการการรับรู้ทางเคมีและพฤติกรรมของปลาของภาควิชา Ichthyology คณะชีววิทยา มหาวิทยาลัยแห่งรัฐมอสโก
ผลงานส่วนตัวของผู้เขียน ผู้เขียนมีส่วนร่วมโดยตรงในการตั้งค่า รับ และประมวลผลสื่อทดลอง และตีความผลลัพธ์ที่ได้รับ เธอมีหน้าที่รับผิดชอบในการแก้ปัญหาที่ได้รับมอบหมายทั้งหมด สรุปผล และพิสูจน์ข้อสรุปทางวิทยาศาสตร์
สิ่งพิมพ์ บทบัญญัติหลักของวิทยานิพนธ์มีอยู่ในงานพิมพ์ 15 เรื่อง
โครงสร้างของวิทยานิพนธ์ วิทยานิพนธ์นี้นำเสนอด้วยข้อความพิมพ์ดีด 171 หน้า ประกอบด้วยตาราง 27 ตาราง ตัวเลข 18 รูป และภาคผนวก 11 ภาค ประกอบด้วย คำนำ 5 บท บทสรุป บทสรุป รายการวรรณกรรมที่อ้างอิง และภาคผนวก รายการแหล่งอ้างอิงประกอบด้วย 260 แหล่ง โดย 150 แหล่งเป็นภาษาต่างประเทศ
รับทราบผู้เขียนรู้สึกซาบซึ้งและขอบคุณหัวหน้างานด้านวิทยาศาสตร์ ศาสตราจารย์ ดุษฎีบัณฑิต สาขาวิทยาศาสตร์ชีวภาพ อ.โอ. ขอขอบพระคุณ กัสสุมยาน สำหรับความช่วยเหลืออันล้ำค่าและการสนับสนุนในทุกขั้นตอนของงานและการจัดทำวิทยานิพนธ์ ผู้เขียนรู้สึกขอบคุณอย่างจริงใจต่อเจ้าหน้าที่ภาควิชา Ichthyology คณะชีววิทยา มหาวิทยาลัยแห่งรัฐมอสโก สำหรับความช่วยเหลือและคำแนะนำที่มีให้ในระหว่างการทำงานและรับสื่อการทดลอง: S.S. Sidorov, E.A. Marusov, T.V. Golovkina, G.V. Devitsina รวมถึงนักศึกษาภาควิชา Ichthyology E.N. Dokuchaeva และ A.N. ฉันไม่สุภาพที่ได้รับความช่วยเหลือในการทำการทดลอง ผู้เขียนแสดงออก
ขอขอบคุณอย่างจริงใจต่อเจ้าหน้าที่ของสถาบันชีววิทยาแห่งน่านน้ำภายในประเทศที่ตั้งชื่อตาม บัตรประชาชน ปาปานีนา (IBVV RAS) Yu.V. สลินโก, ไอ.จี. Grechanov และ Yu.V. Chebotareva สำหรับโอกาสในการใช้ทรายแดงรุ่นเยาว์ในการทดลอง ผู้เขียนแสดงความขอบคุณอย่างจริงใจต่อ D.D. Vorontsov สำหรับการพัฒนาโปรแกรมคอมพิวเตอร์ "BH-Fish" และโอกาสที่จะใช้มันในงานของเขา ผู้เขียนขอขอบคุณผู้อำนวยการสถาบันวิทยาศาสตร์ของรัฐบาลกลาง "สถาบันวิจัยนิเวศวิทยาอ่างเก็บน้ำประมง" ศาสตราจารย์สาขาวิทยาศาสตร์กายภาพและคณิตศาสตร์ศาสตราจารย์ V.N. Lopatin สำหรับความช่วยเหลือทางเทคนิค การให้คำปรึกษา และการสนับสนุนทางการเงินในขั้นตอนสุดท้ายของการเตรียมวิทยานิพนธ์เพื่อการป้องกัน
การกำหนดความชอบด้านรสชาติ
พิพิธภัณฑ์สัตว์น้ำแต่ละแห่งมีเครื่องฉีดน้ำเซรามิกที่มีรูพรุนซึ่งเชื่อมต่อกับไมโครคอมเพรสเซอร์ ด้านบนของตู้ปลาถูกปิดด้วยฝาพลาสติกที่มีรูตรงกลางสำหรับป้อนเม็ดทดลองและให้อาหารปลา ผนังด้านหลังและด้านข้างของถังทำจากพลาสติกทึบแสงหรือกระดาษสีขาวเพื่อแยกปลาออกจากกันด้วยสายตา ซึ่งผู้ทดลองและสังเกตการณ์ได้มองเห็นปลาและเม็ดเล็กลงในถังได้ชัดเจน ผ่านผนังใสด้านหน้าถัง มีการดำเนินการเปลี่ยนน้ำบางส่วนในตู้ปลาทุกสัปดาห์ ในระหว่างการทดลอง ปลาจะได้รับอาหารจนอิ่มด้วยหนอนเลือดวันละครั้งหลังจากสิ้นสุดการทดลอง
การทดลองเชิงพฤติกรรมเพื่อกำหนดรสนิยมของปลาเริ่มต้นหลังจากที่ผู้ทดลองเคยชินกับสภาพการทดลองโดยสมบูรณ์แล้วเท่านั้น ในตอนท้ายของช่วงปรับตัวให้ชินกับสภาพแวดล้อม การก่อตัวของพฤติกรรมปกติของปลาแสดงออกในการเคลื่อนไหวอย่างอิสระทั่วตู้ปลาทั่วไป ไม่มีความขี้อายหรือซ่อนตัวในระหว่างการเคลื่อนไหวหรือการจัดการที่ดำเนินการโดยผู้ทดลองเมื่อสังเกตปลา ให้อาหารหรือทำความสะอาดตู้ปลา มาถึงตอนนี้ปลาก็เริ่มกินอาหารอย่างเต็มใจและคุ้นเคยกับสภาพใหม่ภายใน 2-3 วัน เพื่อประเมินทัศนคติของปลาต่อรสชาติสิ่งเร้า เราใช้วิธีการโดยอาศัยการลงทะเบียนปฏิกิริยาทดสอบพฤติกรรมของปลาและเม็ดวุ้น-วุ้นเทียมที่มีสารรสชาติที่ทดสอบ
ก่อนที่การทดลองจะเริ่มต้นขึ้น ปลาจะได้รับการฝึกให้จับอาหารของตัวอ่อนไคโรโนมิด (หนอนเลือด) ทีละตัว จากนั้นจึงฝึกเม็ดวุ้น-วุ้นด้วยสารสกัดจากน้ำของหนอนเลือด สัตว์ทดลองเรียนรู้อย่างรวดเร็วในการจับเม็ดเม็ดที่นำเข้ามา โดยปกติแล้วปลาจะอยู่ลึกลงไปในน้ำหรือที่ด้านล่างของตู้ปลาใต้รูบนฝา และหยิบเม็ดที่ให้มาในเกือบ 00% ของการนำเสนอ ตามกฎแล้วเม็ดจะถูกตั้งค่าทันทีที่ลงไปในน้ำหรือหลังจาก 2-5 วินาที ข้อยกเว้นคือ verkhovka ในการทดลองซึ่งจับเม็ดได้หลังจากผ่านไป 30.0 วินาทีเท่านั้น บุคคลที่เตรียมด้วยวิธีนี้ก็คว้าสิ่งที่เสนอให้พวกเขาในเวลาต่อมา
หลังจากเสร็จสิ้นการฝึกอบรม พวกเขาเริ่มการทดลองโดยกินเพื่อตรวจสอบปฏิกิริยาทางพฤติกรรมของปลาต่ออูลูที่นำเข้ามาในพิพิธภัณฑ์สัตว์น้ำ ในการทดลองแต่ละครั้งซึ่งเริ่มต้นด้วยการนำเม็ดหนึ่งเข้าไปในตู้ปลา ตัวชี้วัดต่อไปนี้จะถูกบันทึก: จำนวนการกระทำของการจับเม็ดที่แนะนำจนกระทั่งช่วงเวลากลืนหรือการปฏิเสธครั้งสุดท้าย; 2) ระยะเวลาในการคงตัวของแกรนูล (เป็นวินาที) ที่การตั้งค่าครั้งแรก 3) ระยะเวลาที่ปลาถือเม็ดไว้ในปากตลอดการทดลอง; 4) ความอร่อยของเม็ด ได้แก่ เม็ดที่จับได้นั้นถูกกินหรือถูกปฏิเสธ
ระยะเวลาที่ปลายังคงจับเม็ดไว้ถูกบันทึกโดยใช้นาฬิกาจับเวลาแบบแมนนวลของ Agat ประเภทสรุปผล
ช่วงเวลาของการกลืนเม็ดถูกกำหนดโดยความสมบูรณ์ของการเคลื่อนไหวการเคี้ยวลักษณะเฉพาะของขากรรไกรและการฟื้นฟูการเคลื่อนไหวเป็นจังหวะโดยฝาครอบเหงือก การปฏิเสธเม็ดสุดท้ายโดยปลานั้นตัดสินจากพฤติกรรมของมัน ซึ่งแสดงออกในการที่ปลาเคลื่อนออกจากเม็ดและไม่มีการเข้าใกล้หรือพยายามจับมันซ้ำๆ การทดลองที่ปลาไม่จับเม็ดภายใน 1 นาทีหลังจากนำเข้าไปในตู้ปลาจะไม่ถูกนำมาพิจารณาด้วย เม็ดที่ถูกปลาปฏิเสธหรือไม่ได้จับมา (กรณีที่หายากมาก) จะถูกเอาออกจากตู้ปลาทันทีหลังจากสิ้นสุดการทดลอง
สุ่มนำเสนอเม็ดที่มีการกระตุ้นรสชาติต่างกันตามลำดับและสลับกับเม็ดที่มีสารสกัดจากตัวอ่อนไคโรโนมิดที่ความเข้มข้นข้างต้นและด้วยเม็ดควบคุม ช่วงเวลาระหว่างการทดลองกับผู้ทดลองคนเดียวกันคืออย่างน้อย 10-15 นาที จำนวนการทดลองกับแกรนูลประเภทเดียวกันกับแต่ละการทดลองมีความใกล้เคียงกันหรือเหมือนกัน
การศึกษาพลวัตของการตอบสนองต่อรสชาติเชิงพฤติกรรม ในการทดลองชุดพิเศษ ได้ทำการศึกษาพลวัตของการตอบสนองต่อรสชาติเชิงพฤติกรรม เพื่อจุดประสงค์นี้ ระยะเวลาในการจับเม็ด (U) ติดต่อกัน และระยะเวลาระหว่างการจับเม็ด (I) ตลอดจนระยะเวลาตั้งแต่ตกเม็ดลงน้ำจนกระทั่งปลาจับ (I0 ) ได้รับการบันทึกสำหรับปลาที่เลี้ยงในตู้ปลา เพื่อจุดประสงค์เหล่านี้จึงใช้โปรแกรมคอมพิวเตอร์ "BH-Fish" ซึ่งทำให้สามารถกำหนดระยะเวลาของช่วงเวลาเหล่านี้ได้ด้วยความแม่นยำ 0.1 วินาที การลงทะเบียนเหตุการณ์เหล่านี้ (การจับและการปฏิเสธตามลำดับระหว่างการทดลองแยกต่างหาก) ดำเนินการโดยการกดปุ่มที่เกี่ยวข้องบนแป้นพิมพ์หรือเมาส์ของคอมพิวเตอร์ ระยะเวลาของการทดลองทั้งหมด รวมถึงช่วงเวลาแต่ละช่วง (ช่วงการเก็บรักษาเม็ดและช่วงระหว่างการเก็บรักษา) จะถูกบันทึกไว้ในไฟล์แยกต่างหาก จากนั้นข้อมูลเหล่านี้จะถูกนำมาใช้สำหรับการประมวลผลทางสถิติ
กรดอะมิโนอิสระ
ไม่พบสิ่งกระตุ้นที่กำหนดระหว่างกรดอะมิโนและสารแต่งกลิ่นรสแบบดั้งเดิม กรดอะมิโนส่วนใหญ่ -12 จาก 21 ทำให้การบริโภคเม็ดเพิ่มขึ้นอย่างมีนัยสำคัญ (ตารางที่ 6) ซีสเตอีนมีรสชาติที่น่าดึงดูดที่สุดสำหรับเทนช์ซึ่งมีอยู่ในเม็ดทำให้ปลาเกือบ 100% บริโภค ประสิทธิภาพของกรดอะมิโนกระตุ้นอื่นๆ ต่ำกว่ามาก เวลาการเก็บรักษาเฉลี่ยของเม็ดที่มีกรดอะมิโนต่างกันแตกต่างกันอย่างมาก (จาก 2-3 วินาทีถึง 23 วินาที) และมีความสัมพันธ์อย่างใกล้ชิดกับความอร่อย (ตารางที่ 7) ความแปรผันของจำนวนเฉลี่ยของการจับเม็ดในการทดลองกับกรดอะมิโนต่ำกว่า (จาก 1.0 เป็น 2.1) ไม่มีการเปิดเผยความสัมพันธ์ที่สำคัญของตัวบ่งชี้นี้กับพารามิเตอร์อื่น ๆ ของการตอบสนองต่อรสชาติ เม็ดที่มีซิสเตอีนที่น่าดึงดูดมากถูกกลืนโดยปลาส่วนใหญ่หลังจากการจับครั้งแรก (ในการทดลอง 144 ครั้งจาก 146 ครั้งที่กลืนเม็ดเข้าไป) การกักเก็บเม็ดปลาด้วยสารนี้นานกว่ากรดอะมิโนอื่น ๆ 3-5 เท่าหรือมากกว่า (ตารางที่ 8) . โดยเฉลี่ยแล้ว ปลาจะเก็บอาหารเม็ดที่มีกรดอะมิโนที่น่าดึงดูดน้อยกว่าหรือไม่แยแสไว้ในช่วงเวลาที่สั้นกว่า ในขณะที่การปฏิเสธและการจับอาหารเม็ดใหม่เกิดขึ้นบ่อยกว่ามากในระหว่างการทดลอง ตัวอย่างเช่น ในการทดลองบางอย่าง ปลาจะจับเม็ดที่มีซีรีน อาร์จินีน กรดแอสปาร์ติก กรดกลูตามิก หรือไทโรซีนมากถึง 7-8 ครั้งหรือมากกว่านั้น การเปรียบเทียบการบริโภคปลาเม็ดกับออส-ฟีนิลอะลานีนและพี-ฟีนิลอะลานีนไม่พบความแตกต่างที่มีนัยสำคัญทางสถิติ (p <0.05)
เนื่องจากซิสเทอีนมีฤทธิ์กระตุ้นสูง จึงมีการทดลองเพื่อหาค่าขีดจำกัดความเข้มข้นของกรดอะมิโนนี้ เป็นที่ยอมรับกันว่าเมื่อความเข้มข้นลดลง ความน่าดึงดูดของรสชาติของเม็ดก็จะลดลงตามธรรมชาติ เช่นเดียวกับตัวบ่งชี้อื่นๆ ส่วนใหญ่ของการตอบสนองต่อรสชาติ ที่ความเข้มข้น 0.01 M คุณสมบัติที่น่าสนใจของซิสเตอีนจะไม่ปรากฏอีกต่อไป ไม่มีความแตกต่างอย่างมีนัยสำคัญที่เกี่ยวข้องกับการควบคุมพารามิเตอร์การตอบสนองอื่น ๆ (จำนวนการจับและระยะเวลาการเก็บรักษาเม็ดโดยปลา) จะสังเกตได้ที่ ความเข้มข้นของซิสเทอีน 0.001 M (ตารางที่ 8)
กรดอินทรีย์
กรดอินทรีย์ส่วนใหญ่ - 17 จาก 19 - มีรสชาติที่น่าดึงดูดสำหรับเทนช์ ผลการกระตุ้นของกรด 7 ชนิด ได้แก่ มาลิก, α-คีโตกลูตาริก, ออกซาลิก, ทาร์ทาริก, มาลิก, ซิตริกและมาโลนิกนั้นสูงที่สุด (ระดับการบริโภคของเม็ดมากกว่า 70%) เม็ดที่อยู่กับพวกมันถูกกลืนส่วนใหญ่หลังจากการจับครั้งแรกและค้างอยู่ในช่องปากเป็นเวลานาน กรดเพียงสองตัวเท่านั้นที่ไม่ส่งผลกระทบที่เห็นได้ชัดเจนต่อการบริโภคปลาเม็ด - อะซิติกและโชลิค
บันทึก. , - ระดับนัยสำคัญตาม /? 0.05, 0.01, 0.001 ก่อนเฉือนครั้งแรก - ตอบสนองต่อรสชาติของกรดอะมิโนอิสระ ระหว่างสแลช - สำหรับกรดอินทรีย์ หลังจากเฉือนครั้งที่สอง - รวมกรดอะมิโนอิสระ กรดอินทรีย์ และสารแต่งกลิ่นรสดั้งเดิม
สำหรับกรดอินทรีย์ โดยทั่วไป มีความสัมพันธ์ที่ชัดเจนระหว่างระดับการบริโภคเม็ด จำนวนครั้งในการจับเม็ด และระยะเวลาที่ปลากักไว้ (ตารางที่ 9)
การทดลองเพื่อกำหนดระดับความไวต่อรสชาติของเทนช์ต่อกรดอินทรีย์ดำเนินการโดยใช้ตัวอย่างของกรดมาลิกซึ่งเป็นกรดที่มีประสิทธิภาพมากที่สุดชนิดหนึ่ง เป็นที่ยอมรับแล้วว่าผลการกระตุ้นของกรดมาลิกปรากฏในช่วงความเข้มข้น 0.1-0.01 M เมื่อความเข้มข้นลดลงตามลำดับความสำคัญอื่น (0.001 M) ผลจะไม่ปรากฏ เมื่อความเข้มข้นลดลง ปริมาณการใช้แกรนูลและระยะเวลาที่ปลากักไว้จะลดลงตามธรรมชาติ และจำนวนการจับในการทดลองจะเพิ่มขึ้น (ตารางที่ 8)
ในการทดลองกับมัสตาร์ด มีการใช้สารแต่งกลิ่นรสดั้งเดิม กรดอะมิโนอิสระ และกรดอินทรีย์ นอกจากนี้ยังกำหนดระดับความไวของความขม ขม ต่อสารแต่งกลิ่นที่มีคุณสมบัติยับยั้งด้วย
รสชาติคลาสสิก
พบว่าในบรรดาสารปรุงแต่งรสแบบดั้งเดิมมีเพียงกรดซิตริกเท่านั้นที่สามารถระคายเคืองได้อย่างมีประสิทธิผลสำหรับรสขมทำให้การบริโภคเม็ดลดลงอย่างรวดเร็ว (ตารางที่ 10) ซูโครส แคลเซียมคลอไรด์ และโซเดียมคลอไรด์เป็นตัวกระตุ้นรสชาติที่ไม่แยแส เม็ดที่มีสารสกัดจากหนอนเลือดถูกรับประทานอย่างง่ายดายที่สุด
จำนวนเหตุการณ์การจับเม็ดโดยเฉลี่ยในการทดลองอยู่ระหว่าง 1.3-1.9 (สารสกัดจากหนอนเลือดและสิ่งเร้าที่ไม่แยแส) ถึง 2.3-2.1 (การควบคุมและกรดซิตริก)
เวลาที่ปลากักเก็บเม็ดจะยิ่งใหญ่ที่สุดในการทดลองกับสารสกัด bloodworm เท่านั้น และสั้นที่สุดในการทดลองกับกรดซิตริก ในกรณีอื่นๆ ทั้งหมด ระยะเวลาของการเก็บเม็ดไม่แตกต่างจากกลุ่มควบคุมอย่างมีนัยสำคัญ เวลาที่ปลาเก็บเม็ดที่ไม่สวยงาม (ที่มีกรดซิตริก) ไว้นั้นต่ำเป็นประวัติการณ์ - 0.6 วินาทีเมื่อจับครั้งแรก และ 1.4 วินาทีตลอดการทดลองทั้งหมด
รสชาติคลาสสิก
ในบรรดากรดอะมิโนอิสระพบกรดอะมิโนกระตุ้น 8 ชนิดและกรดอะมิโนยับยั้ง 1 ตัว - ไอโซลิวซีนระดับการบริโภคของแกรนูลซึ่งเท่ากับ 0 นั่นคือไม่มีแกรนูลใดของประเภทนี้ที่ถูกกลืนโดยปลาหลังจากการตั้งค่า (ตารางที่ 22). กรดอะมิโนที่เหลืออีก 12 ชนิดไม่มีผลกระทบอย่างมีนัยสำคัญต่อการบริโภคอาหารเม็ดและทำหน้าที่เป็นสารที่ไม่แยแสสำหรับปลา เม็ดที่มีอะลานีนมีรสชาติที่น่าดึงดูดที่สุด (59.3%) ระดับการบริโภคซึ่งสูงกว่ารสชาติของเม็ดที่มีสารสกัดจากหนอนเลือด (57.1%) นอกจากนี้ตามระดับของผลการกระตุ้นที่ลดลงกรดอะมิโนจะถูกจัดเรียงตามลำดับต่อไปนี้: ไกลซีน, ซีสเตอีน, กลูตามีน, ไลซีน, แอสพาราจีน, วาลีน, ทรีโอนีน จำนวนชุดเม็ดเล็กแตกต่างกันไปในช่วงตั้งแต่ 1.5 ถึง 2.8 การทดลองกับแกรนูลที่มีกรดอะมิโนกระตุ้น ได้แก่ อะลานีน ไกลซีน แอสพาราจีน และทรีโอนีน อาร์จินีนของกรดอะมิโนที่ไม่แยแส กรดแอสปาร์ติก และนอร์วาลีน และไอโซลิวซีนของกรดอะมิโนที่ยับยั้ง ได้รับการแยกแยะด้วยการจับซ้ำจำนวนเล็กน้อยอย่างมีนัยสำคัญ ระยะเวลาการเก็บรักษาเม็ดมีความสัมพันธ์อย่างมากกับระดับความอร่อย (rs=0.96; pO.001) (ตารางที่ 23) เวลากักเก็บที่สั้นที่สุดคือในการทดลองกับไอโซลิวซีน ซึ่งไม่น่าดึงดูดสำหรับปลาทรายแดง (1.0 วินาที - หลังจากการจับครั้งแรก และ 1.7 วินาทีในระหว่างการทดลองทั้งหมด) และกับกรดอะมิโนที่ไม่แยแสบางชนิด เช่น นอร์วาลีน ไทโรซีน ทริปโตเฟน และเมไทโอนีน
คุณลักษณะเฉพาะของการตอบสนองต่อรสชาติของทรายแดงคือการจับเม็ดทรายซ้ำๆ เป็นจำนวนมาก โดยสามารถจับได้ถึง 22-25 ครั้งในการทดลองบางอย่าง เช่นเดียวกับการคงสภาพของเม็ดทรายไว้ในระยะยาว (ในการทดลองบางอย่างที่ใช้เวลานานกว่า 1 นาที)
เพื่อชี้แจงการเปลี่ยนแปลงของการตอบสนองพฤติกรรมรสชาติของเทนช์ จึงมีการใช้แกรนูลที่มีอะลานีน การเลือกกรดอะมิโนนี้เกิดจากการที่ในบรรดาสารที่ทดสอบทั้งหมดในการทดลองกับอะลานีนพบว่ามีการจับกุมเม็ดเล็ก ๆ โดยเฉลี่ยค่อนข้างมาก - 1.6 (ตารางที่ 6) สิ่งสำคัญคือก่อนที่จะกลืนหรือปฏิเสธเม็ดในที่สุด ปลาจะเก็บมันไว้ในช่องปากเป็นเวลานาน นอกจากนี้ การเลือกอะลานีนยังถูกกำหนดโดยระดับการใช้เม็ดเล็กเกือบ 50% อีกด้วย ซึ่งทำให้สามารถวิเคราะห์ไดนามิกของการตอบสนองต่อรสชาติแยกกันสำหรับการทดลองที่จบลงด้วยการกลืนกิน (การทดลอง SG) หรือการปฏิเสธเม็ด (การทดลอง OG) . จากมุมมองของระเบียบวิธีที่สำคัญเหล่านี้ คุณลักษณะของการตอบสนองของเทนช์ต่อเม็ดที่มีอะลานีนช่วยอำนวยความสะดวกในการแก้ปัญหาในการอธิบายพลวัตของการตอบสนองรสชาติเชิงพฤติกรรมของปลาอย่างมีนัยสำคัญ จากปลาทดลอง 15 ตัว มีการเลือกตัวบุคคล 2 ตัวสำหรับการทดลอง ซึ่งพารามิเตอร์การตอบสนองของแต่ละตัวสะท้อนถึงค่าเฉลี่ยของกลุ่มได้ใกล้เคียงที่สุด
ปริมาณการใช้เม็ดอะลานีนในการทดลองชุดนี้คือ 54% และระยะเวลาเฉลี่ยของการทดลองคือ 12.4 วินาที ในการทดลองส่วนใหญ่ พบว่ามีการจับ 1-2 เม็ด จำนวนการจับสูงสุดคือ 6 (รูปที่ 2) ระยะเวลาเฉลี่ยของการทดลองขึ้นอยู่กับจำนวนการจับซ้ำของแกรนูล (รูปที่ 3) และแปรผันจาก 5.7 วินาที ในกรณีของการจับหนึ่งครั้ง จนถึง 33.1 วินาที ด้วยการจับ 6 ครั้ง (ตารางที่ 25) โดยเฉลี่ย ระยะเวลาการเก็บรักษาของเม็ดสั้นกว่า (2.72 วินาที) กว่าช่วงระหว่างการจับ (4.52 วินาที) อย่างมีนัยสำคัญ ที่ยาวที่สุดคือการยึดเกาะครั้งแรกของแกรนูลและช่วงระหว่างการจับที่ 1 และ 2 (ตารางที่ 25)
ระยะเวลาติดต่อกันของการตอบสนองต่อรสชาติเชิงพฤติกรรมของเทนช์ต่อเม็ดที่มีแอล-อะลานีนคือ 0.1 M. U - U6 - ช่วงแรกและช่วงต่อมาของการเก็บรักษาเม็ดตามลำดับ HJ-J - I5-6 - ตามลำดับ ช่วงเวลาระหว่างครั้งแรกและครั้งที่สอง และการตั้งค่าภายหลังของแกรนูล การกำหนดที่เหลือเป็นไปตามรูปที่ 3
ผลลัพธ์ที่คล้ายกันยังได้รับเมื่อวิเคราะห์พลวัตของการแสดงออกของการตอบสนองพฤติกรรมการรับรสของปลาแยกกันในการทดลองที่จบลงด้วยการกลืนหรือปฏิเสธอาหารอัดเม็ด - การยึดหลังจากการจับครั้งแรกจะยาวที่สุดกว่าครั้งต่อ ๆ ไปทั้งหมด และระยะเวลาของการจับ ลดลงเมื่อมีการจับเม็ดแต่ละครั้ง ช่วงเวลาระหว่างการจับลดลงอย่างต่อเนื่องในการทดลอง OG และยังคงอยู่ที่ระดับเดียวกันโดยประมาณในการทดลอง SG ระยะเวลาในการตอบสนองต่อรสชาติจะนานขึ้นในการทดลอง 3G (ตารางที่ 25 รูปที่ 2 และ 3)
เม็ดซิสเทอีน
ปลามีลักษณะเฉพาะด้วยการปรับตัวของสายพันธุ์ที่เกี่ยวข้องกับโภชนาการที่หลากหลายสูงมาก (Nikolsky, 1956, 1971, 1974; Wootton, 1998) ธรรมชาติและองค์ประกอบของอาหารที่ปลาบริโภค ความกว้างและความแปรปรวนของสิ่งมีชีวิตที่ใช้เป็นอาหาร วิธีการได้รับอาหาร จังหวะการให้อาหาร ฯลฯ แตกต่างกันไป เชื่อกันว่าตัวบ่งชี้เหล่านี้ปลามีความเหนือกว่าสัตว์มีกระดูกสันหลังอื่น ๆ อย่างมีนัยสำคัญ (Nikolsky, 1974) ในเครือข่ายทางโภชนาการที่มีการจัดการอย่างซับซ้อนและมีหลายองค์ประกอบซึ่งก่อตัวขึ้นในระบบนิเวศทางน้ำและทำให้เกิดความมั่นคงและความยั่งยืน ปลามักเป็นผู้บริโภคในลำดับสูงสุด (Biro, 1998) การศึกษาโภชนาการปลา ลักษณะของพฤติกรรมการให้อาหาร ฐานประสาทสัมผัสในการค้นหา การตรวจจับ และการเลือกวัตถุอาหาร อิทธิพลต่อประสิทธิภาพการให้อาหารของสภาพแวดล้อมภายนอกต่างๆ และสถานะแรงจูงใจภายในและสถานะทางสรีรวิทยาของปลาเป็นของ การวิจัยด้านวิทยาวิทยาแบบดั้งเดิม (Shorygin, 1952; Nikolsky, 1956 ; Gaevskaya, 1956; Manteuffel et al., 1965; Fortunatova, Popova, 1973; Ivlev, 1977; Hiatt, 1983; Mikheev, 2006) คำถามเหล่านี้ยังคงได้รับการพัฒนาอย่างแข็งขันในปัจจุบัน โดยให้ความสนใจเป็นพิเศษกับปัญหาในการอธิบายกลไกและรูปแบบที่เป็นรากฐานของการดำเนินการของปลาโดยมีลักษณะทางชีววิทยาที่ซับซ้อนที่เกี่ยวข้องกับโภชนาการและพฤติกรรมการให้อาหาร - นิเวศวิทยา สรีรวิทยา และจริยธรรม และอื่นๆ (Hart, Gill, 1992; Pavlov, Kasumyan, 1990, 1998; Kasumyan, 1997; Osse et al., 1997; Kasumyan, Doving, 2003) การพัฒนาปัญหาพื้นฐานที่สำคัญเหล่านี้เป็นเรื่องที่น่าสนใจอย่างยิ่งในการแก้ปัญหาเร่งด่วนหลายประการของการเพาะเลี้ยงสัตว์น้ำและการประมงสมัยใหม่ เช่น การเพิ่มประสิทธิภาพและการสร้างวิธีการและเทคโนโลยีใหม่ในการให้อาหาร การพัฒนาอาหารสัตว์เทียม การเพิ่มความสามารถในการจับเครื่องมือประมง เป็นต้น (จ็อบลิง, 1994; จ็อบลิง และคณะ, 1995) ในการสนับสนุนทางประสาทสัมผัสของพฤติกรรมการให้อาหาร - ลำดับการกระทำที่ซับซ้อนตั้งแต่การรับสัญญาณอาหารและการค้นหาเหยื่อที่เป็นไปได้ ไปจนถึงการตรวจจับ การประเมินเบื้องต้น การจับ การประมวลผลภายในช่องปาก และการกลืนกินหรือการปฏิเสธ ระบบเซ็นเซอร์เคมีมีบทบาทสำคัญ การมีส่วนร่วมของการรับรู้รสชาติประกอบด้วยการตรวจสอบคุณภาพของเหยื่อที่จับโดยปลา การประเมินคุณสมบัติด้านรสชาติ และการปฏิบัติตามความต้องการทางโภชนาการของแต่ละบุคคล (Kasumyan และ Doving, 2003)
เป็นที่ทราบกันดีว่าปฏิกิริยาทางพฤติกรรมของสัตว์ส่วนใหญ่รวมทั้งปลานั้นเกิดขึ้นจากข้อมูลที่ได้รับผ่านช่องทางประสาทสัมผัสต่างๆ พร้อมๆ กัน ทั้งระบบรับสัมผัสทางเคมีหลัก - การดมกลิ่นและการรับรส - สามารถมีส่วนร่วมในการก่อตัวของการตอบสนองเชิงพฤติกรรมของปลาต่อเม็ดที่มีสารเคมี ในการก่อตัวของการตอบสนองของปลาต่อเม็ด บทบาทของระบบเคมีเซ็นเซอร์ที่ 3 ซึ่งเป็นความไวต่อสารเคมีทั่วไปนั้นดูเหมือนจะมีน้อย เนื่องจากตามการศึกษาทางอิเล็กโทรสรีรวิทยาที่ดำเนินการเป็นพิเศษได้แสดงให้เห็นว่า ระบบนี้มีช่วงของสิ่งเร้าที่มีประสิทธิภาพที่จำกัดอย่างยิ่ง (Kotrschal et al. , 1996) คำตอบสำหรับคำถามที่ว่าระบบรับกลิ่นหรือการรับลมระบบใดที่รับผิดชอบต่อการตอบสนองเชิงพฤติกรรมต่ออาหารเม็ดที่เราบันทึกไว้ในปลาเป็นสิ่งสำคัญโดยพื้นฐาน การทดลองกับปลาที่ไม่มีออสเมตเป็นวิธีดั้งเดิมในการอธิบายบทบาทของกลิ่นในการตอบสนองทางพฤติกรรมของปลาต่อสัญญาณทางเคมีประเภทต่างๆ (Kasumyan และ Pashchenko, 1982; Liley et al., 1993; Kasumyan และ Devitsina, 1997 และอื่นๆ อีกมากมาย) การวิเคราะห์เปรียบเทียบของการทดลองเชิงพฤติกรรมกับบุคคลที่ไม่มีการควบคุมและควบคุม (ไม่บุบสลาย) ของปลาคาร์พและสร้อยซึ่งดำเนินการโดยใช้วิธีการที่เหมือนกับของเราแสดงให้เห็นว่าปลาที่ปราศจากความไวในการดมกลิ่นไม่ได้นำไปสู่การเปลี่ยนแปลงปฏิกิริยาต่อเม็ดกับสาร ในปลาที่ไม่มีออสเมตและไม่บุบสลาย ความเข้มข้นเกณฑ์ของสารที่ใส่เข้าไปในแกรนูลก็ตรงกันเช่นกัน ข้อมูลเหล่านี้เป็นเหตุให้สรุปได้ว่าระบบรับกลิ่นไม่เกี่ยวข้องกับการสนับสนุนทางประสาทสัมผัสของความแตกต่างที่สังเกตได้ในการตอบสนองเชิงพฤติกรรมของปลาทดลองต่อเม็ดที่มีสารต่างกัน และยังว่าธรรมชาติและความรุนแรงของการตอบสนองเหล่านี้ได้มาจากการรับรู้รสชาติในช่องปากด้วย (Kasumyan, Morsi, 1996; Kasumyan, Sidorov, 2005)
ผลลัพธ์ของเราแสดงให้เห็นว่าปลาไซปรินิดที่ศึกษาสามารถจดจำและตอบสนองต่อเม็ดที่มีสารแต่งกลิ่นได้แตกต่างกัน เช่น สารแต่งกลิ่นรสดั้งเดิม กรดอะมิโนอิสระ และกรดอินทรีย์ ในบรรดาการตอบสนองรสชาติเชิงพฤติกรรมของปลาเหล่านี้สามารถสังเกตคุณสมบัติทั่วไปอย่างหนึ่ง: ซูโครสในทุกสายพันธุ์ไม่มีผลกระทบอย่างมีนัยสำคัญต่อระดับการบริโภคเม็ดนั่นคือ เป็นตัวกระตุ้นการรับรสที่ไม่แยแส (ตารางที่ 26 รูปที่ 15) คุณลักษณะเฉพาะของการตอบสนองต่อรสชาติต่อสิ่งเร้ารสคลาสสิกคือกรดซิตริกมีคุณสมบัติด้านรสชาติที่น่าดึงดูดใจอย่างมากสำหรับทรายแดงและเทนช์ และมีรสชาติที่น่ารังเกียจอย่างมากสำหรับปลาคาร์พ crucian สีทองและ verkhovka โซเดียมคลอไรด์ทำให้ระดับการบริโภคแกรนูลในเทนช์และทรายแดงเพิ่มขึ้นอย่างมีนัยสำคัญและสำหรับสายพันธุ์อื่นมันเป็นสารที่ไม่แยแส แคลเซียมคลอไรด์ทำหน้าที่เป็นตัวกระตุ้นรสชาติที่มีประสิทธิภาพสำหรับปลาเทนช์ทรายแดงและปลาคาร์พ crucian สีทอง ทัศนคติของ verkhovka ต่อรสชาติของแคลเซียมคลอไรด์นั้นเป็นลบและในความขมขื่นก็ไม่แยแส
วิทยานิพนธ์
อิซาวา, โอลก้า มิคาอิลอฟนา
ระดับการศึกษา:
ผู้สมัครสาขาวิทยาศาสตร์ชีวภาพ
สถานที่รับวิทยานิพนธ์:
รหัสพิเศษ HAC:
ความชำนาญพิเศษ:
วิทยา
เลขหน้า:
บทที่ 1 การทบทวนวรรณกรรม
บทที่ 2 วัสดุและวิธีการ
2.1. วัตถุประสงค์ของการวิจัย
2.2. เงื่อนไขและการเตรียมปลาเพื่อการทดลอง
2.3. การกำหนดความชอบด้านรสชาติ
2.4. ศึกษาพลวัตของการตอบสนองรสนิยมทางพฤติกรรม
2.5. การเตรียมแกรนูลทดลอง
2.6. ลักษณะทั่วไปของวัสดุ
บทที่ 3 การตอบสนองทางพฤติกรรมของปลา Cypird ต่อรสชาติคลาสสิก กรดอะมิโนอิสระ และกรดอินทรีย์
3.1.1. รสชาติคลาสสิก
3.1.2. กรดอะมิโนอิสระ
3.1.3. กรดอินทรีย์
3.2. กอร์ชัค
3.2.1. รสชาติคลาสสิก
3.2.2. กรดอะมิโนอิสระ
3.2.3. กรดอินทรีย์
3.3. เวอร์คอฟกา
3.3.1. รสชาติคลาสสิก
3.3.2. กรดอะมิโนอิสระ
3.4. ปลาคาร์ปทอง
3.4.1. รสชาติคลาสสิก
3.4.2. กรดอะมิโนอิสระ
3.5. ทรายแดง
3.5.1. รสชาติคลาสสิก
3.5.2. กรดอะมิโนอิสระ
บทที่ 4 พลวัตของการสำแดงพฤติกรรมของ GASTE
คำตอบด้วยปลาคาร์ป
4.2.1. เม็ดซิสเทอีน
4.2.2. เม็ดกลูตามีน
4.3. กอร์ชัค
บทที่ 5 การอภิปรายผล 86 บทสรุป 123 บทสรุป 127 ข้อมูลอ้างอิงที่อ้างถึง 127 ภาคผนวก
การแนะนำวิทยานิพนธ์ (ส่วนหนึ่งของบทคัดย่อ) ในหัวข้อ “ความชอบและพฤติกรรมรสชาติของปลาคาร์พ”
ความเกี่ยวข้องของหัวข้อ พฤติกรรมการให้อาหารเป็นรากฐานของหน้าที่ที่สำคัญที่สุดของอาหารปลา ซึ่งก็คือโภชนาการ ซึ่งมีความสำคัญอย่างยิ่งต่อทั้งต่อบุคคลและประชากรและสายพันธุ์โดยรวม บทบาทนำในการสนับสนุนทางประสาทสัมผัสในช่วงสุดท้ายของพฤติกรรมการให้อาหารปลานั้นมีบทบาทโดยการรับรสชาติ ซึ่งรับประกันการประเมินคุณสมบัติด้านรสชาติของเหยื่อและสอดคล้องกับความต้องการทางโภชนาการของปลาตลอดจนการบริโภคอาหารที่เพียงพอ รายการ (Atema, 1980; Pavlov, Kasumyan, 1990, 1998; Kasumyan, 1997 ) จนกระทั่งเมื่อไม่นานมานี้ มีการศึกษาเกี่ยวกับระบบการรับรสของปลาจำนวนมากเพื่อชี้แจงลักษณะทางสัณฐานวิทยาและภูมิประเทศของปุ่มรับรสเป็นหลัก โครงสร้างพิเศษและปกคลุมด้วยเส้นการศึกษาการจัดโครงสร้างทางสัณฐานวิทยาของส่วนกลางของระบบรสชาติ (Zuwala, Jakubowski, 1993; Reutter, 1992; Jakubowski, Zuwala, 2000; Vasilevskaya, 1974) การศึกษาจำนวนมากเกี่ยวกับคุณสมบัติเชิงหน้าที่ของระบบรสชาติของปลาดำเนินการโดยใช้วิธีอิเล็กโทรสรีรวิทยาเป็นหลัก (Marui และ Caprio, 1992; Jones, 1990; Sutterlin, 1975) อย่างไรก็ตามเมื่อเร็ว ๆ นี้ทิศทางอื่นในการศึกษาการรับรู้รสชาติได้รับการพัฒนาอย่างรวดเร็วและมีประสิทธิผลมาก: นี่เป็นวิธีทดสอบปฏิกิริยาเชิงพฤติกรรมด้วยความช่วยเหลือซึ่งคุณสามารถประเมินความชอบด้านรสชาติของปลาจำนวนมากได้ ปริมาณข้อมูลการทดลองที่สะสมในช่วงไม่กี่ปีที่ผ่านมาทำให้มีแนวคิดที่ชัดเจนเกี่ยวกับรูปแบบทั่วไปและคุณลักษณะเฉพาะของความสัมพันธ์ระหว่างปลากับสิ่งเร้าในรสชาติ ความเหมือนและความแตกต่างในสเปกตรัมอาหารของสารปรุงแต่งรสที่มีประสิทธิภาพในปลาทุกวัยและเป็นระบบ ตำแหน่ง และยังแสดงความเหมือนและความแตกต่างระหว่างการรับรสภายนอกและในช่องปากด้วย (Kasumyan, 1997; Kasumyan, D/aving, 2003)
แต่จนถึงขณะนี้คำถามเกี่ยวกับการมีอยู่ของความคล้ายคลึงและ/หรือความแตกต่างในความชอบด้านรสชาติยังคงมีการศึกษาที่ไม่ดีนัก ที่เกี่ยวข้องอย่างใกล้ชิดชนิดของปลา ไม่มีข้อมูลเกี่ยวกับลักษณะทางสรีรวิทยาของการแสดงการตอบสนองต่อพฤติกรรมการรับรสในปลา ไม่มีข้อมูลเกี่ยวกับอิทธิพลของวิถีชีวิตของปลาไซปรินิดสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิดต่อการแสดงออกขององค์ประกอบต่าง ๆ ของการตอบสนองเชิงพฤติกรรมต่อสิ่งเร้ารสชาติต่างๆ
การศึกษาประเด็นเหล่านี้และประเด็นสำคัญอื่นๆ เกี่ยวกับการรับรู้รสชาติของปลามีความสำคัญทางทฤษฎี นอกเหนือจากการปฏิบัติจริงแล้ว ทำให้สามารถชี้แจงบทบาทของระบบประสาทสัมผัสนี้ในการรับรองการเลือกให้อาหารปลาด้วยรายการอาหารที่เพียงพอและในการรักษาสภาวะสมดุล ความรู้เกี่ยวกับรูปแบบของความไวต่อรสชาติของปลาลักษณะการตอบสนองต่อสารปรุงแต่งประเภทต่างๆมีความเกี่ยวข้องกับความเป็นไปได้ในการแก้ปัญหาเฉพาะที่ของการเพาะเลี้ยงสัตว์น้ำสมัยใหม่เช่นการค้นหาสารกระตุ้นทางโภชนาการทางเคมีที่มีประสิทธิภาพสูงการปรับปรุงที่มีอยู่และการสร้าง อาหารเทียมชนิดใหม่ การพัฒนาเทคโนโลยีชีวภาพสำหรับการเลี้ยงปลา ตลอดจนเหยื่อเคมีเทียมและเหยื่อปลา
เป้าหมายของการทำงาน เพื่อศึกษาความชอบด้านรสชาติ ความไวต่อสิ่งเร้ารสชาติประเภทต่างๆ และลักษณะของพฤติกรรมการรับรสในปลาสายพันธุ์ที่เกี่ยวข้องกันอย่างใกล้ชิด (โดยใช้ตัวอย่างปลาในตระกูลปลาคาร์พ Cyprinidae)
วัตถุประสงค์ของการศึกษาประกอบด้วย:
เพื่อเปรียบเทียบความชอบด้านรสชาติของสารปรุงแต่งรสดั้งเดิมและกรดอะมิโนอิสระในปลาไซปรินิด
เพื่อค้นหาความชอบด้านรสชาติของกรดอินทรีย์ในปลาคาร์พ
ตรวจสอบความสัมพันธ์ระหว่างรสชาติที่น่าดึงดูดของสารกับคุณสมบัติทางโครงสร้างและเคมีกายภาพบางประการ
กำหนดระดับความไวต่อรสชาติของปลาคาร์พต่อสารปรุงแต่งที่มีคุณสมบัติกระตุ้นและยับยั้ง
เพื่อศึกษาพฤติกรรมการกินของปลาคาร์พ โครงสร้าง พลวัต และลักษณะขององค์ประกอบหลักของการตอบสนองพฤติกรรมการกินของปลาคาร์พ
ความแปลกใหม่ทางวิทยาศาสตร์ ความแปลกใหม่ทางวิทยาศาสตร์ของงานวิทยานิพนธ์นี้อยู่ที่การขยายความเข้าใจลักษณะการทำงานของระบบรสชาติในปลาสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิด และอธิบายลักษณะของการตอบสนองพฤติกรรมรสชาติที่พวกมันแสดงออกมา งานนี้เป็นครั้งแรกที่กำหนดรสนิยมของปลาคาร์พ 5 สายพันธุ์สำหรับสารปรุงแต่งรสคลาสสิกและกรดอะมิโนอิสระ และปลา 2 สายพันธุ์สำหรับกรดอินทรีย์
เป็นครั้งแรกที่มีการกำหนดระดับความไวต่อรสชาติของปลาคาร์พ 3 สายพันธุ์ต่อสารที่มีคุณสมบัติด้านรสชาติเชิงบวก รวมถึงสารที่ทำให้เกิดการตอบสนองต่อรสชาติเชิงลบ (สารยับยั้ง) แสดงให้เห็นว่าไม่มีความเชื่อมโยงที่เหมือนกันกับปลาคาร์พระหว่างระดับความน่าดึงดูดของรสชาติของสารและคุณสมบัติทางเคมีกายภาพ (น้ำหนักโมเลกุล ค่า pH ของสารละลาย จำนวนกลุ่มฟังก์ชัน) เป็นครั้งแรกที่ใช้โปรแกรมคอมพิวเตอร์ต้นฉบับ "BH-P13" เพื่อศึกษาโครงสร้างของการตอบสนองรสชาติเชิงพฤติกรรมของปลา พลวัตของการแสดงออกได้รับการชี้แจง และระยะเวลาของการกระทำตามพฤติกรรมแต่ละอย่างของการตอบสนองต่อรสชาติถูกกำหนดแยกต่างหากสำหรับ การทดลองที่สิ้นสุดด้วยการกินหรือการปฏิเสธวัตถุอาหารเทียม (อาหารเม็ด)
ความสำคัญเชิงปฏิบัติของงาน ผลการศึกษานี้สามารถนำไปใช้ในการปฏิบัติงานด้านการเพาะเลี้ยงสัตว์น้ำและการประมงในการค้นหาสารกระตุ้นเคมีอาหารที่มีประสิทธิภาพสูง ในการปรับปรุงและพัฒนาเหยื่อและสิ่งที่แนบมาเทียม ในการเตรียมและปรับปรุงสูตรอาหารเทียมเพื่อเพิ่มความน่ารับประทานสำหรับ ปลา. ผลลัพธ์ที่ได้รับจากพลวัตของการตอบสนองพฤติกรรมรสชาติทำให้เรามีความคิดที่ชัดเจนยิ่งขึ้นเกี่ยวกับลักษณะของการแสดงออกของพฤติกรรมการให้อาหารที่จัดอย่างซับซ้อนโดยปลาในระยะสุดท้ายและวิธีการควบคุมโดยตรงโดยใช้สิ่งกระตุ้นทางเคมี ผลการศึกษานี้ใช้เป็นส่วนหนึ่งของหลักสูตรการบรรยาย "สรีรวิทยาของปลา" ซึ่งมอบให้กับนักศึกษาภาควิชา Ichthyology คณะชีววิทยา มหาวิทยาลัยแห่งรัฐมอสโก บทบัญญัติที่ได้รับการคุ้มครอง
1. ความชอบด้านรสชาติของปลาสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิดซึ่งอยู่ในตระกูลเดียวกัน (ไซปรินิด) มีลักษณะเฉพาะคือมีความเฉพาะเจาะจงของสายพันธุ์สูง
2. ความอร่อยของสารบางชนิดอาจจะเหมือนกันในปลาสายพันธุ์ที่เกี่ยวข้องกันอย่างใกล้ชิด
3. ความเข้มข้นของสารที่มีคุณสมบัติดึงดูดและน่ารังเกียจสำหรับปลาใกล้เคียงกัน
4. พฤติกรรมการกินของปลามีโครงสร้างและไดนามิกที่แน่นอน โดยมีลักษณะเฉพาะและทั่วไปในปลาที่มีวิถีชีวิตและพฤติกรรมการกินอาหารที่แตกต่างกัน และขึ้นอยู่กับคุณสมบัติด้านรสชาติของวัตถุที่เป็นอาหาร
งานนี้ดำเนินการโดยได้รับการสนับสนุนทางการเงินจากมูลนิธิรัสเซียเพื่อการวิจัยขั้นพื้นฐาน (ทุน 04-0448157 และ 07-04-00793) และอยู่ภายใต้กรอบแผนงานเฉพาะเรื่องของสถาบันวิทยาศาสตร์แห่งรัฐ "NIIERV"
การอนุมัติงาน มีการนำเสนอเอกสารวิทยานิพนธ์ในการประชุม All-Russian Symposium " อายุและสรีรวิทยาสิ่งแวดล้อมของปลา"(Borok, IBVV, 1998), การประชุมระหว่างมหาวิทยาลัยนานาชาติ "Lomonosov-98" (Moscow, MSU, 1998), การประชุมระหว่างมหาวิทยาลัยครั้งที่ 2 ที่อุทิศให้กับวันอนุรักษ์พื้นที่ชุ่มน้ำโลก (Rybnoye, 1999), การประชุมทางจริยธรรมนานาชาติครั้งที่ 26 (บังกาลอร์, อินเดีย, 1999) ), การประชุมนานาชาติ "การเชื่อมต่อทางโภชนาการในชุมชนทางน้ำและระบบนิเวศ" (Borok, 2003), การประชุมทางวิทยาศาสตร์ระดับนานาชาติครั้งที่สอง "เทคโนโลยีชีวภาพ - การคุ้มครองสิ่งแวดล้อม" (มอสโก, 2004), การประชุมนานาชาติ " ปัญหาสมัยใหม่ทางสรีรวิทยาและชีวเคมีของสิ่งมีชีวิตในน้ำ"(Petrozavodsk, 2004), การประชุมนานาชาติ "พฤติกรรมของปลา" (Borok, 2005), การประชุมใหญ่ครั้งที่ 9 ของหอดูดาวแห่งรัฐของ Russian Academy of Sciences (Tolyatti, 2006), การประชุมนานาชาติ "ปัญหานิเวศวิทยาประชากรของสัตว์" (Tomsk, 2549), การประชุมนานาชาติครั้งที่ 4 เรื่อง “การสื่อสารกับสัตว์เคมี. ปัญหาพื้นฐาน" (มอสโก, 2549) ที่การประชุมเชิงปฏิบัติการของห้องปฏิบัติการการรับรู้ทางเคมีและพฤติกรรมของปลาของภาควิชา Ichthyology คณะชีววิทยา มหาวิทยาลัยแห่งรัฐมอสโก
ผลงานส่วนตัวของผู้เขียน ผู้เขียนมีส่วนร่วมโดยตรงในการตั้งค่า รับ และประมวลผลสื่อทดลอง และตีความผลลัพธ์ที่ได้รับ เธอมีหน้าที่รับผิดชอบในการแก้ปัญหาที่ได้รับมอบหมายทั้งหมด สรุปผล และพิสูจน์ข้อสรุปทางวิทยาศาสตร์
สิ่งพิมพ์ บทบัญญัติหลักของวิทยานิพนธ์มีอยู่ในงานพิมพ์ 15 เรื่อง
โครงสร้างของวิทยานิพนธ์ วิทยานิพนธ์นี้นำเสนอด้วยข้อความพิมพ์ดีด 171 หน้า ประกอบด้วยตาราง 27 ตาราง ตัวเลข 18 รูป และภาคผนวก 11 ภาค ประกอบด้วย คำนำ 5 บท บทสรุป บทสรุป รายการวรรณกรรมที่อ้างอิง และภาคผนวก รายการแหล่งอ้างอิงประกอบด้วย 260 แหล่ง โดย 150 แหล่งเป็นภาษาต่างประเทศ
บทสรุปของวิทยานิพนธ์ ในหัวข้อ "Ichthyology", Isaeva, Olga Mikhailovna
1. ปลาคาร์พสายพันธุ์ที่ศึกษามีความไวต่อรสชาติที่ชัดเจนต่อสารเคมีประเภทต่างๆ - กรดอะมิโนฟรี, กรดอินทรีย์, สารแต่งกลิ่นแบบดั้งเดิม
2. สเปกตรัมรสชาติของกรดอะมิโนอิสระ กรดอินทรีย์ และสารแต่งกลิ่นแบบดั้งเดิมในปลาไซปรินิดมีความแตกต่างกันในด้านความกว้าง องค์ประกอบ และประสิทธิผลสัมพัทธ์ของสาร ในกรณีส่วนใหญ่ ไม่พบความสัมพันธ์ที่มีนัยสำคัญระหว่างสเปกตรัมรสชาติของปลาคาร์พ ซึ่งยืนยันระดับสูงของความจำเพาะของสายพันธุ์ของความชอบในรสชาติของปลา รวมถึง ที่เกี่ยวข้องอย่างใกล้ชิดสายพันธุ์.
3. ความชอบรสชาติของปลาคาร์พในแต่ละสารอาจจะเหมือนกันหรือคล้ายกันก็ได้ ระดับความไวต่อรสชาติต่อสารที่มีคุณสมบัติด้านรสชาติที่น่าดึงดูดหรือน่ารังเกียจสำหรับปลาไม่แตกต่างกันอย่างมีนัยสำคัญ
4. ไอโซเมอร์เชิงโครงสร้างมีความอร่อยที่แตกต่างกันสำหรับปลา และทำให้เกิดการตอบสนองต่อรสชาติที่แตกต่างกันในลักษณะเชิงปริมาณ ไม่พบการเชื่อมโยงทั่วไปกับปลาคาร์พระหว่างระดับความน่าดึงดูดของรสชาติของสารและคุณสมบัติทางโครงสร้างและคุณสมบัติทางเคมีกายภาพ (น้ำหนักโมเลกุล ค่า pH ของสารละลาย จำนวนหมู่ฟังก์ชัน ฯลฯ)
5. ลักษณะเฉพาะของพฤติกรรมการกินของปลาคือการจับและปฏิเสธวัตถุอาหารซ้ำ ๆ ก่อนกลืนหรือปฏิเสธการบริโภคในขั้นสุดท้าย จำนวนการจับซ้ำ ระยะเวลาในการจับวัตถุ ตลอดจนเวลาทั้งหมดที่ใช้ในการทดสอบอาหารปลา ขึ้นอยู่กับรสชาติของวัตถุอาหาร ตลอดจนวิถีชีวิตและพฤติกรรมการให้อาหารของปลา
6. พฤติกรรมการกินเกิดขึ้นในลักษณะเดียวกันในปลาต่างสายพันธุ์ ในกรณีที่ปลากลืนเหยื่อหรือปฏิเสธที่จะกินในที่สุด พฤติกรรมการกินจะแตกต่างกันไปตามระยะเวลาในการเก็บวัตถุอาหารอย่างต่อเนื่อง และช่วงเวลาระหว่างการจับและในการเปลี่ยนแปลงของพารามิเตอร์เหล่านี้
บทสรุป
ผลลัพธ์ที่ได้จากการทำงานนี้ยืนยันถึงความจำเพาะของสเปกตรัมรสชาติในปลาในระดับสูง รวมถึงสายพันธุ์ที่เกี่ยวข้องอย่างใกล้ชิดด้วย เอกลักษณ์เฉพาะของสเปกตรัมรสชาตินั้นสัมพันธ์กับกลุ่มสารประกอบที่ศึกษาทั้งหมด - สารปรุงแต่งรสคลาสสิก, กรดอะมิโนอิสระ, กรดอินทรีย์ ในเวลาเดียวกัน ปลาที่มีความเกี่ยวข้องกันอย่างใกล้ชิดอาจมีทัศนคติต่อรสชาติของสารบางชนิดที่คล้ายคลึงกันหรือเหมือนกัน ซึ่งบ่งบอกถึงความคล้ายคลึงบางประการในความชอบของพวกมัน ไม่ต้องสงสัยเลยว่าจำเป็นต้องมีการวิจัยเพิ่มเติมเพื่อกำหนดข้อสรุปที่เข้มงวดมากขึ้นเกี่ยวกับความสัมพันธ์ระหว่างความชอบด้านรสชาติและการเชื่อมโยงอย่างเป็นระบบของปลา การทำงานต่อเนื่องในทิศทางนี้และการเพิ่มจำนวนพันธุ์ปลาที่ศึกษาจะช่วยค้นหาคำตอบสำหรับคำถามอื่นๆ ที่ยังตอบได้ไม่เพียงพอเกี่ยวกับการรับรู้รสชาติของปลา โดยเฉพาะเกี่ยวกับความสัมพันธ์ระหว่างการเลือกรสชาติของปลากับวิถีชีวิตและรูปแบบการให้อาหาร .
ในงานที่เกี่ยวข้องกับการรับรู้รสชาติของปลานั้นให้ความสนใจอย่างมากในการค้นหาลักษณะทางโครงสร้างหรือเคมีฟิสิกส์ของสารเคมีที่สามารถอธิบายหรือสัมพันธ์กับคุณสมบัติด้านรสชาติของปลาได้ ผลการวิจัยพบว่าปลามีระดับความชอบด้านรสชาติที่แตกต่างกันสำหรับสารที่มีคุณสมบัติทางโครงสร้างของโมเลกุลหรือคุณสมบัติต่างกัน อย่างไรก็ตาม ไม่มีการระบุความเชื่อมโยงที่เหมือนกันกับปลาระหว่างลักษณะโครงสร้างและเคมีกายภาพของสารกับความน่าดึงดูดของรสชาติ ซึ่งอธิบายได้ด้วยสเปกตรัมรสชาติเฉพาะสายพันธุ์
เป็นครั้งแรกที่ได้รับข้อมูลเกี่ยวกับโครงสร้างพฤติกรรมการรับรสและการเปลี่ยนแปลงของพฤติกรรมในปลา การกลืนกินหรือการปฏิเสธเม็ดในขั้นสุดท้ายเกิดขึ้นในปลาหลังจากการปฏิเสธหลายครั้งและการจับเม็ดซ้ำหลายครั้ง ยิ่งการจับเม็ดละเอียดซ้ำๆ กันมากเท่าใด ปลาก็จะใช้เวลานานขึ้นในการประเมินคุณภาพรสชาติของเหยื่อเท่านั้น สิ่งนี้ใช้ได้กับการทดลองที่สิ้นสุดด้วยการกินอาหารอัดเม็ดและการทดลองซึ่งท้ายที่สุดแล้วปลาก็ปฏิเสธอาหารเม็ดนั้น ระยะเวลาในการตอบสนองต่อรสชาติในกรณีหลังนั้นสั้นกว่ามาก สาเหตุหลักมาจากระยะเวลาการกักเก็บเม็ดยาในช่องปากสั้นลง ระยะเวลาในการกักเก็บเม็ดเล็กอย่างรวดเร็วและเป็นธรรมชาติจะลดลงตามการตั้งค่าแต่ละครั้ง ในขณะที่ช่วงเวลาระหว่างการตั้งค่าจะเปลี่ยนไปอย่างมีนัยสำคัญน้อยลง การเปลี่ยนแปลงของการตอบสนองต่อรสชาติจะแตกต่างกันไปในรสขม ทรายแดง และเทนช์ อาจเนื่องมาจากความแตกต่างในลักษณะและกลยุทธ์การให้อาหารปลาและวิถีชีวิตของพวกมัน ปลาที่อาศัยอยู่ในน้ำนิ่งและกินโดยตัวแทนของสัตว์ในท้องทะเลเป็นหลัก (ปลาทรายแดง, ปลาเทนช์) มีแนวโน้มที่จะวิเคราะห์วัตถุรับรสเป็นเวลานานเนื่องจากมีโอกาสเพิ่มขึ้นที่อนุภาคดินที่กินไม่ได้จะเข้าไปในช่องปากและจำเป็นต้องแยกวัตถุอาหารออก . ปลาเหล่านี้ยังจับเม็ดอาหารซ้ำหลายครั้งอีกด้วย
รูปแบบของการรับรู้รสชาติในปลา คุณลักษณะเฉพาะของการตอบสนองของปลาต่อสารปรุงแต่งชนิดต่างๆ และพลวัตของการแสดงการตอบสนองของพฤติกรรมด้านรสชาติ ถือเป็นความสนใจในทางปฏิบัติที่สำคัญ และสามารถนำมาใช้ในการแก้ปัญหาต่างๆ ในการประมงและการเพาะเลี้ยงสัตว์น้ำ การศึกษาที่ดำเนินการแสดงให้เห็นถึงคำมั่นสัญญาในการค้นหาและสร้างสารกระตุ้นและสารยับยั้งที่มีประสิทธิภาพสูงสำหรับปลา และทำหน้าที่เป็นพื้นฐานทางชีวภาพสำหรับการพัฒนาวิธีการควบคุมพฤติกรรมการให้อาหารปลาโดยใช้สิ่งกระตุ้นรสชาติ ผลลัพธ์ที่เราได้รับสามารถใช้เพื่อเพิ่มความน่ารับประทานของอาหารสัตว์ เหยื่อตกปลา และเหยื่อ เพื่อดำเนินการแก้ไขสูตรโดยการนำสารพิเศษที่มีฤทธิ์กระตุ้นสูง หรือโดยการแยกส่วนประกอบที่มีสารประกอบยับยั้งออกจากส่วนประกอบ สิ่งนี้จะไม่เพียงแต่ลดการสูญเสียอาหารเทียมโดยตรงเท่านั้น แต่ยังรับประกันว่าการเปลี่ยนอาหารสำหรับการเจริญเติบโตของปลามีประสิทธิภาพมากขึ้นอีกด้วย เป็นที่ทราบกันดีว่าการบริโภคอาหารที่น่าสนใจซึ่งมีการรับสัมผัสทางเคมีนั้นมาพร้อมกับปลาที่มีการหลั่งเอนไซม์ย่อยอาหารที่รุนแรงมากขึ้น (Takesla และ Takp, 1992)
รายการอ้างอิงสำหรับการวิจัยวิทยานิพนธ์ ผู้สมัครสาขาวิทยาศาสตร์ชีวภาพ Isaeva, Olga Mikhailovna, 2550
1. อันดริยาเชฟ เอ.พี. พ.ศ. 2487 บทบาทของอวัยวะรับความรู้สึกในการหาอาหารในทะเล Burbot // Journal. ชีวการแพทย์ทั่วไป ต.5 ลำดับที่ 2. หน้า 123-127.
2. อันดริยาเชฟ เอ.พี. 2498. บทบาทของอวัยวะรับความรู้สึกในการหาอาหารในปลา // Tr. การประชุมวิธีการศึกษาแหล่งอาหารและโภชนาการของปลา อ.: สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต. หน้า 135-142.
3. Aristovskaya G.V. พ.ศ. 2478 ในประเด็นโภชนาการของปลาโวลก้า - คามาบางชนิด // Tr. สาขาตาตาร์ของ VNIORH ต. 2.
4. อาร์โนลด์ที่ 2 พ.ศ. 2445 การสังเกตการให้อาหารปลาในบางพื้นที่ของ Valdai Upland // " ผู้ประกาศข่าวอุตสาหกรรมประมง" ลำดับที่ 1.
5. เบลี เอ็น.ดี. 2499. ชีววิทยาและการเพาะพันธุ์ทรายแดง. เคียฟ: จาก Academy of Sciences ของ SSR ของยูเครน หน้า 45-54.
6. โบโบรฟ ยู.พี. พ.ศ. 2511 โภชนาการและการเจริญเติบโตของปลาคาร์พหญ้าในสภาพฟาร์มบ่อในเขตภาคกลางของ RSFSR // งานวิจัยใหม่เกี่ยวกับนิเวศวิทยาและการผสมพันธุ์ สัตว์กินพืชปลา อ.: วิทยาศาสตร์. หน้า 106-115.
7. โบโดรวา เอ็น.วี. พ.ศ. 2505 ตัวรับความรู้สึกทางเคมีของทรายแดง // ปัญหาทางวิทยาวิทยา ท.2 ฉบับที่ 4 (25) หน้า 48-54.
8. โบโดรวา เอ็น.วี. พ.ศ. 2508 การจัดโครงสร้างของตัวรับกลิ่นของปลา // ในการรวบรวม ไบโอนิค อ.: วิทยาศาสตร์. หน้า 48-69.
9. โบรุตสกี้ อี.วี. พ.ศ. 2493 วัสดุเกี่ยวกับโภชนาการของปลาคาร์พเงินอามูร์ (Hypophthalmichthys molitrix Val.) // การดำเนินการของอามูร์ วิทยาการสำรวจ พ.ศ. 2488-2492 ต. 1.
10. วาซิเลฟสกายา เอ็น.อี. พ.ศ. 2517 ว่าด้วยการรับสารเคมีโดยไม่รับกลิ่นในปลา // ในคอลเลกชัน: ลักษณะพื้นฐานของพฤติกรรมและทิศทางของปลา อ.: วิทยาศาสตร์. หน้า 36-56.
11. เวอริจิน บี.วี. พ.ศ. 2504 ผลการดำเนินงานเพื่อปรับสภาพปลาที่กินพืชเป็นอาหารจากตะวันออกไกลและมาตรการในการพัฒนาและศึกษาเพิ่มเติมในพื้นที่ใหม่ // คำถามเกี่ยวกับ Ichthyology ท.1 ฉบับ. 4 (21) หน้า 640-649.
12. เกฟสกายา เอ็น.เอส. พ.ศ. 2499 ภารกิจหลักในการศึกษาแหล่งอาหารและโภชนาการปลาในแง่มุมพื้นฐานที่สำคัญที่สุดของการประมง // การประชุมเรื่องวิธีการศึกษาแหล่งอาหารและโภชนาการปลา อ.: สำนักพิมพ์ของสถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต หน้า 6-20.
13. Gdovsky P.A., Gremyachikh V.A., Nepomnyashchikh V.A. พ.ศ. 2537 ผลของภาวะ anosmia ต่อระดับน้ำตาลในเลือดและพฤติกรรมการสำรวจของปลาคาร์พ Cyprinus carpio ต่อหน้าจุดสังเกตที่มองเห็นได้ // Zh. วิวัฒนาการ ชีวเคมีและฟิสิกส์ ต.30. ลำดับที่ 6 ป.746-752.
14. เกอร์ซา ไอ.ไอ. 2524. การส่องสว่างและพฤติกรรมของปลา. อ.: วิทยาศาสตร์. 163 หน้า
15. แกรนด์ดิเลฟสกายา-เดกซ์บัค M.J1. พ.ศ. 2504 คุณสมบัติหลักของสัตว์ด้านล่างและโภชนาการปลาของอ่างเก็บน้ำ Kama (พ.ศ. 2498-2502) // Tr. อูราล แผนก กอสNIORH. ลำดับที่ 5.
16. เดวิทซินา จี.วี. พ.ศ. 2540 ในประเด็นการสนับสนุนทางเคมีและสัมผัสของพฤติกรรมการให้อาหารปลาคอดในทะเลสีขาว // ประเด็น. วิทยา ต.37 ลำดับที่ 1 น.94-100.
17. เดวิทซินา จี.วี. 2541. การพัฒนาอวัยวะรับสารเคมีในการสร้างยีนของหอกทั่วไป Esox lucius II ฉบับ. วิทยา ต. 38 ลำดับ 4 หน้า 537-547
18. เดวิทซินา จี.วี. 2547. ระบบเคมีเซ็นเซอร์ของปลา: การจัดโครงสร้าง-หน้าที่และปฏิสัมพันธ์ // บทคัดย่อของผู้เขียน. โรค . วิทยาศาสตรดุษฎีบัณฑิต วิทยาศาสตร์ ม.44 น.
19. เดวิทซินา จี.วี. 2548. โครงสร้างของเครื่องรับรสในช่องปากของปลาที่เกี่ยวข้องกับพฤติกรรมการให้อาหารโดยเฉพาะ // การประชุมนานาชาติเรื่อง "พฤติกรรมของปลา". โบร็อก หน้า 131-138.
20. Devitsyna G.V., Gadzhieva A.R. 2539 พลวัตของการพัฒนาทางสัณฐานวิทยาของระบบรสชาติในช่วงต้นของการกำเนิดของตัวแทนสองคนของปลาสเตอร์เจียน Acipenser nudiventris และ A. persicus // คำถาม วิทยา ต.36.ลำดับ5.ป.674-686.
21. Devitsyna G.V., Kazhlaev A.A. 2538. ระบบเคมีเซ็นเซอร์และการสร้างสัณฐานวิทยาแบบเฮเทอโรโครนิกในปลาสเตอร์เจียนรุ่นเยาว์ตอนต้น ชีวฟิสิกส์ ต. 40. ประเด็น. 1.ส. 146-150.
22. เดวิทสินา จี.วี., กัสสุมยาน เอ.โอ. 2543. ปฏิสัมพันธ์ส่วนกลางของระบบเคมีบำบัดในปลาสเตอร์เจียน // ระบบประสาทสัมผัส. ต.14.ข้อ2.หน้า107-117
23. ดิมิเทรียวา อี.เอช. 2500 การวิเคราะห์ทางสัณฐานวิทยาของปลาคาร์พ crucian สองสายพันธุ์ // Tr. สถาบันสัณฐานวิทยาสัตว์ตั้งชื่อตาม เอ.เอ็น. เซเวิร์ตโซวา ต.16.
24. Dmitrieva T.M., Moskaleva T.M. 1984. อิเล็กโทรสรีรวิทยาลักษณะของความสำคัญเชิงหน้าที่ของการรับเคมีบำบัดโดยไม่รับกลิ่นในปลา // Tr. 1 ทั้งหมด การประชุมทางสรีรวิทยาทางประสาทสัมผัสของปลา มูร์มันสค์. หน้า 49-51.
25. ดอมราเชฟ พี.เอฟ., ปราฟดิน ไอ.เอฟ. พ.ศ. 2505 ปลาในทะเลสาบอิลเมนและแม่น้ำ Volkhov และความสำคัญด้านการประมง // Mat. จากการวิจัยพบว่า ร. Volkhov และแอ่งน้ำ เลนินกราด ฉบับที่ X
26. ดราโกมิรอฟ เอ็น.ไอ. 2497. การพัฒนาตัวรับผิวหนังที่ด้านล่างของศีรษะในตัวอ่อนของปลาสเตอร์เจียนที่เปลี่ยนไปสู่วิถีชีวิตหน้าดิน // Dokl. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต.97 ลำดับที่ 1 น. 173-176.
27. เอโกรอฟ เอ.จี. พ.ศ. 2531 ปลาในอ่างเก็บน้ำทางตอนใต้ของไซบีเรียตะวันออก (คล้ายปลาคาร์พ คล้ายปลาค็อด คล้ายคอน) อีร์คุตสค์: จากมหาวิทยาลัยอีร์คุตสค์ 328 หน้า
28. ซิเทเนวา ที.เอส. พ.ศ. 2523 การให้อาหารทรายแดงใน biotopes ต่าง ๆ ของอ่างเก็บน้ำ Rybinsk // Inf. แถลงการณ์ " ชีววิทยาของน่านน้ำภายในประเทศ" เลนินกราด: วิทยาศาสตร์ หมายเลข 46. หน้า 26-30.
29. จูคอฟ พี.ไอ. พ.ศ. 2508 ปลาแห่งเบลารุส มินสค์: สำนักพิมพ์ "วิทยาศาสตร์และเทคโนโลยี" 415 หน้า
30. ซาโดริน เอ.เอ., ซูเอฟ ไอ.วี., วีเชโกรอดต์เซฟ เอ.เอ. 2547 Verkhovka (.Leucaspius delineatus (Heckel)) สายพันธุ์ที่รุกรานในแหล่งน้ำของดินแดนครัสโนยาสค์ // คำถามเกี่ยวกับวิทยาวิทยา ลำดับที่ 1. หน้า 75-79.
31. ซเวเรวา อี.วี. 2535. ความไวต่อรสชาติของปลาแซลมอนบางชนิดและอิทธิพลของค่า pH ของน้ำต่ำที่มีต่อมัน // มอสโก, มหาวิทยาลัยแห่งรัฐมอสโก, คณะชีววิทยา, ภาควิชา วิทยา งานบัณฑิต. ป.86
32. ซเวเรวา โอ.เอส., คูชิน่า อี.เอส., ออสโตรมอฟ เอ็น.เอ. พ.ศ. 2496 ปลาและการประมงบริเวณตอนกลางและตอนล่างของ Pechora อ.: จากสถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต1. หน้า 131-139.
33. อิฟเลฟ VS. 2520. ทดลองนิเวศวิทยาการให้อาหารปลา. เคียฟ: วิทยาศาสตร์ ความคิด 272 หน้า
34. คาสซิล วี.จี. พ.ศ. 2515 รสชาติ // สรีรวิทยาของระบบประสาทสัมผัส. 4.2. ล.: วิทยาศาสตร์. ป.562-606.
35. คาสซิล วี.จี. 2533. พฤติกรรมการกินในการถ่ายทอดทางพันธุกรรม. ล.: วิทยาศาสตร์. 220 หน้า
36. เกษมยาน อ.โอ. 2533. สรีรวิทยาทางประสาทสัมผัสของปลาทะเล. ด้านระเบียบวิธี ไม่แยแส ป.57.
37. เกษมยาน อ.โอ. 2534. กลไกทางประสาทสัมผัสเพื่อความน่าเชื่อถือของการดำเนินการ. เคมีสื่อสารในปลา // ใน: ปัญหาการสื่อสารทางเคมีของสัตว์. ม.วิทยาศาสตร์ หน้า 263-270.
38. เกษมยาน อ.โอ. 2540. การรับรู้รสชาติและพฤติกรรมการให้อาหารปลา // ประเด็น. วิทยา ต.37. ลำดับที่ 1. หน้า 78-93.
39. เกษมยาน อ.โอ. 2548. ความชอบด้านรสชาติและพฤติกรรมการรับรสของปลา // การประชุมนานาชาติเรื่อง "พฤติกรรมของปลา". โบร็อก หน้า 225-227.
40. กสุมยาน อ.โอ., เดวิทสินา จี.วี. พ.ศ. 2540 อิทธิพลของการกีดกันดมกลิ่นต่อความไวของสารเคมีและสถานะของต่อมรับรสของปลาสเตอร์เจียน // ประเด็น. วิทยา ต. 37 ลำดับ 6 หน้า 823-835
41. Kasumyan A.O., Kazhlaev A.A. พ.ศ. 2536 การตอบสนองเชิงพฤติกรรมของเด็กและเยาวชนตอนต้นของปลาสเตอร์เจียนไซบีเรียน Acipenser baeri และปลาสเตอร์เจียน stellate A. stellatus (Acipenseridae) ต่อสารที่ทำให้เกิดความรู้สึกรับรสประเภทหลัก // Vopr. วิทยา ต. 33 ลำดับ 3 หน้า 427-443
42. คาสุมยาน เอ.โอ., มารุซอฟ อี.เอ. 2545. การตอบสนองเชิงพฤติกรรมของปลาซิว Phoxinus phoxinus (Cyprinidae) ต่อสัญญาณทางเคมีภายใต้สภาวะปกติและหลังภาวะ anosmia เฉียบพลันและเรื้อรัง // ปัญหา วิทยา ต.42.ลำดับ5.ป.684-696.
43. คาสุมยาน เอ.โอ., มารุซอฟ อี.เอ. 2546. การตอบสนองเชิงพฤติกรรมของปลาซิวทั่วไปที่ไม่บุบสลายและมี anosmated เรื้อรัง Phoxinus phoxinus (Cyprinidae) ต่อกรดอะมิโนอิสระ // ปัญหา. วิทยา ต. 43 ลำดับที่ 4 หน้า 528-539
44. คาสุมยาน เอ.โอ., มารุซอฟ อี.เอ. 2548. แบบแผนของปฏิกิริยาพฤติกรรมอาหารในปลาตามปกติและหลังภาวะ anosmia เฉียบพลันและเรื้อรัง // การประชุมนานาชาติเรื่อง "พฤติกรรมของปลา". โบร็อก หน้า 227-232.
45. Kasumyan A.O., Morey A.M.H., 2539. ความไวต่อรสชาติของปลาคาร์พ // ปัญหา. วิทยา ต.36.ลำดับ3.ป.386-399.
46. Kasumyan A.O., Morey A.M.Kh., 1997. ความชอบด้านรสชาติของสารปรุงแต่งรสคลาสสิกของปลาคาร์พหญ้าวัยอ่อน Ctenopharyngodon idella (Cyprinidae, Pisces) เลี้ยงด้วยอาหารที่แตกต่างกัน // Dokl หนึ่ง. ต. 357 ลำดับ 2 หน้า 284-286
47. Kasumyan A.O., Morey A.M.H., 1998. ผลของโลหะหนักต่อกิจกรรมการให้อาหารและการตอบสนองพฤติกรรมการรับรสของปลาคาร์พ Cyprinus carpio 1. ทองแดง แคดเมียม สังกะสี และตะกั่ว // ปัญหา. วิทยา ต. 38 ลำดับ 3 หน้า 393-409
48. Kasumyan A.O., Nikolaeva E.V. 2540. รสนิยมความชอบของปลาหางนกยูง Poecilia reticulata II Issue. วิทยา ต. 37 ลำดับ 5 หน้า 696-703
49. Kasumyan A.O., Pashchenko N.I. พ.ศ. 2525 การประเมินบทบาทของกลิ่นในปฏิกิริยาป้องกันของปลาคาร์พหญ้า Ctenopharyngodon idella (Val.) (Cyprinidae) ต่อฟีโรโมนปลุก // คำถามเกี่ยวกับ Ichthyology ต. 22. ประเด็น 2. หน้า 303-307.
50. Kasumyan A.O., Ponomarev V.Yu. พ.ศ. 2529 การศึกษาพฤติกรรมของปลาเซเบริช Brachidanio rerio Hamilton-Buchanan (Cypriniformes, Cyprinidae) ภายใต้อิทธิพลของสัญญาณอาหารเคมีธรรมชาติ // ประเด็น วิทยา ต.26. ฉบับที่ 4. หน้า 665-673.
51. คาสุมยาน เอ.โอ., ซิโดรอฟ เอส.เอส. พ.ศ. 2535 ความไวต่อรสชาติของปลาแซลมอนชุม ออนโครินคัสชุมต่อสิ่งเร้ารสชาติหลักและกรดอะมิโน // ระบบประสาทสัมผัส. ต. 6. ข้อ 3. หน้า 100-103.
52. คาสุมยาน เอ.โอ., ซิโดรอฟ เอส.เอส. พ.ศ. 2536 การตอบสนองเชิงพฤติกรรมของปลาเทราท์แคสเปียนรุ่นเยาว์ Salmo trutta caspius Kessler ต่อสารปรุงแต่งประเภทหลัก // แถลงการณ์ของมหาวิทยาลัยแห่งรัฐมอสโก เซอร์ 16. ชีววิทยา. ลำดับที่ 2. ป.48-54.
53. คาสุมยาน เอ.โอ., ซิโดรอฟ เอส.เอส. 1994ก. การเปรียบเทียบการตอบสนองต่อรสชาติภายในช่องปากและภายนอกช่องปากต่อกรดอะมิโนอิสระในปลาสเตอร์เจียนสามสายพันธุ์ในสกุล Acipenserll Biophysics ต.39 เลขที่. 3. หน้า 526-529.
54. คาสุมยาน เอ.โอ., ซิโดรอฟ เอส.เอส. 25379. รสชาติคุณสมบัติของกรดอะมิโนอิสระสำหรับปลาเทราท์แคสเปียนวัยอ่อน Salmo trutta caspius Kessler // คำถาม วิทยา ต. 34 ลำดับ 6 หน้า 831-838
55. Kasumyan A.O., Sidorov S.S., 1995. การวิเคราะห์เปรียบเทียบการตอบสนองรสชาติของปลาเทราต์เด็กและเยาวชน Salmo trutta trutta จากประชากรของทะเลแคสเปียน, ทะเลบอลติกและทะเลสีขาว // Dokl รศ. ต. 343 ลำดับ 3 หน้า 417-419
56. Kasumyan A.O., Sidorov S.S., 2001. ความไวต่อรสชาติของถ่านทะเลสาบเด็กและเยาวชน Salvelinus namaycush (Salmonidaé) II ปัญหาการประมง ภาคผนวก 1 หน้า 121-125
57. คาสุมยาน เอ.โอ., ซิโดรอฟ เอส.เอส. 2548ก. รสนิยมความชอบของปลาเทราต์สีน้ำตาล Salmo trutta จากประชากรสามกลุ่มที่อยู่ห่างไกลตามภูมิศาสตร์ Vopr วิทยา ต. 45 ลำดับที่ 1 หน้า 117-130
58. คาสุมยาน เอ.โอ., ซิโดรอฟ เอส.เอส. 25489 ผลของการอดอาหารต่อการตอบสนองพฤติกรรมการรับรสของปลาคาร์พ // สื่อการประชุมนานาชาติเรื่อง “พฤติกรรมปลา”. โบร็อก หน้า 237-240.
59. เกษมยาน A.O., Taufik J.P. 2536. พฤติกรรมตอบสนองของปลาสเตอร์เจียนวัยอ่อน (Acipenseridae) ต่อกรดอะมิโน // Vopr. วิทยา ต. 33. ลำดับ 5. หน้า 691-700.
60. Kasumyan A.O., Morey A.M.Kh., Sidorov S.S. พ.ศ. 2536 ความไวต่อรสชาติของปลาคาร์พ Cyprynus carpió ต่อสารที่ทำให้เกิดความรู้สึกรับรสประเภทหลัก // Dokl. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต. 330 ลำดับ 6 หน้า 792-793
61. Kasumyan A.O., Sidorov S.S., Pashchenko N.H. 2536. ผลของอุณหภูมิของน้ำต่อความไวต่อรสชาติของ Acipenser stellatus ปลาสเตอร์เจียนรุ่นเยาว์ต่อกรดอะมิโนอิสระ // Dokl. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต.331. N2. ป.248-250.
62. Kasumyan A.O., Taufik L.R., Protsenko Yu.V. 2534. ความไวในการรับกลิ่นและการรับรสของปลาสเตอร์เจียนรุ่นเยาว์ต่อกรดอะมิโน // พื้นฐานทางชีวภาพของการเพาะพันธุ์ปลาสเตอร์เจียนทางอุตสาหกรรม. อ.: วนิโร. หน้า 37-53.
63. Kasumyan A.O., Kazhlaev A.A., Sidorov S.S., Pashchenko N.N. พ.ศ. 2534 ความน่าดึงดูดใจในการรับกลิ่นและรสชาติของส่วนประกอบอาหารเทียมสำหรับปลาสเตอร์เจียนเด็กและเยาวชน // การประมง ลำดับ 12. หน้า 53-55
64. Kasumyan A.O., Sidorov S.S., Pashchenko N.I., Nemchinov A.B. 2535 ความไวต่อรสชาติภายนอกและในช่องปากของปลาสเตอร์เจียนรัสเซียวัยเยาว์ Acipenser gueldenstaedti ต่อกรดอะมิโน // Dokl. สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ต. 322 หมายเลข 1.ส. 193-195.
65. คุปชินสกี้ บี.เอส. พ.ศ. 2530 ปลาทรายแดงในแหล่งน้ำของลุ่มน้ำไบคาล-อังการา อีร์คุตสค์: จากมหาวิทยาลัยอีร์คุตสค์ 144 น.
66. คิริลลอฟ เอฟ.เอ็น. พ.ศ. 2515 ปลาแห่งยาคุเตีย อ.: วิทยาศาสตร์. 360 วิ
67. Lebedev V.D., Spanovskaya V.D. 1983. วงศ์ Cyprinidae IM. ชีวิตของสัตว์ ต.4. ปลา. ป.228-272.
68. ลินด์เบิร์ก G.U. พ.ศ. 2490 ปลาตัวอ่อนของเอเชียกลาง อ.: จากสถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต หน้า 78-90.
69. ลูปาเชวา แอล.ไอ. พ.ศ. 2510 การให้อาหารปลาคาร์พหญ้าในระยะแรกของการพัฒนา // Rybn. ครัวเรือน เคียฟ ฉบับที่ 3. หน้า 102-104.
70. Manteifel B.P., Girsa I.I., Lescheva T.S., Pavlov D.S. พ.ศ. 2508 จังหวะโภชนาการและการเคลื่อนไหวของปลานักล่าในน้ำจืดบางชนิดในแต่ละวัน // โภชนาการของปลานักล่าและความสัมพันธ์กับสิ่งมีชีวิตในอาหาร อ.: วิทยาศาสตร์. ป.3-81.
71. มิคาอิโลวา อี.เอส., คาซุมยาน เอ.โอ. 2548. การเปรียบเทียบการตอบสนองพฤติกรรมการรับรสของ Stickleback สามหนามจากประชากรที่อยู่ห่างไกลทางภูมิศาสตร์หลายแห่ง // การดำเนินการของการประชุมนานาชาติเรื่อง "พฤติกรรมของปลา". โบร็อก หน้า 336-340.
72. Morey A.M.H. , 1995. ความไวต่อรสชาติของปลาคาร์พและการเปลี่ยนแปลงภายใต้อิทธิพลของโลหะหนัก / M.: Moscow State University, วิทยานิพนธ์ สำหรับการสมัครงาน เอ่อ ขั้นตอน, Ph.D., 172 p.
73. นิโคลสกี้ จี.วี. พ.ศ. 2499 ปลาในลุ่มน้ำอามูร์ ผลลัพธ์ของการสำรวจ ichthyological ของอามูร์ในปี พ.ศ. 2488-2492 อ.: จากสถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต 551 ส
74. นิโคลสกี้ จี.วี. 2514. วิทยาเฉพาะทาง. ม.: มัธยมปลาย. 471 น.
75. นิโคลสกี้ จี.วี. 2517. นิเวศวิทยาของปลา. ม.: มัธยมปลาย. 174 น.
76. Nikolsky G.V., Gromicheva N.A., Morozova G.I., Pikuleva V.A. พ.ศ. 2490 ปลาในแอ่ง Pechora ตอนบน // ปลาในแอ่ง Pechora ตอนบน อ.: จาก-ใน
77. มอพ. วัสดุความรู้เกี่ยวกับสัตว์และพืชในสหภาพโซเวียต ตอนใหม่. ภาควิชาสัตววิทยา. ฉบับที่ 6 (XXI) หน้า 5-209.
78. Nikolsky P.D. , Zhdanova N.N. 2502. อิทธิพลของช่วงเวลาน้ำท่วมฟาร์มปลาต่อการอยู่รอดของปลาทรายแดงรุ่นเยาว์ // การประมง. ลำดับที่ 2. หน้า 15-18.
79. Nikolaeva E.V., Kasumyan A.O., 2000. การวิเคราะห์เปรียบเทียบความชอบด้านรสชาติและการตอบสนองต่อพฤติกรรมต่อสิ่งเร้าในรสชาติของปลาหางนกยูงตัวเมียและตัวผู้ Poecilia reticulata I Vopr วิทยา ต.40 ลำดับที่ 4.ป.560-565.
80. Nikolaeva E.V., Kasumyan A.O., 2001. ความชอบด้านรสชาติของปลาลิ้นหมาขั้วโลกรุ่นเยาว์ Liopsetta gîacialis และปลาดุกลาย Anarhichas lupus II ปัญหาการประมง ภาคผนวก 1 หน้า 197-201
81. นิคอนอฟ G.I. 2541 “ เงินมีชีวิต” Ob-Irtysh Tyumen: สำนักพิมพ์ "OFT Design" หน้า 98-180.
82. โอซินอฟ เอ.จี. 2527. ต้นกำเนิดของปลาเทราต์สีน้ำตาลสมัยใหม่ Salmo trutta L. (Salmonidae): ข้อมูลเกี่ยวกับเครื่องหมายของยีนทางชีวเคมี // ฉบับ. วิทยา ต. 24. ประเด็น. 1. น. 11-24.
83. โอซินอฟ เอ.จี., เบอร์นาซ เจ1. พ.ศ. 2539 "แอตแลนติก" และ "ดานูบ" สายวิวัฒนาการกลุ่มปลาเทราท์ Salmo trutta complex: ความแตกต่างทางพันธุกรรม วิวัฒนาการ การป้องกัน // ประเด็น วิทยา ต. 36. ฉบับที่ 6 หน้า 762-786
84. พาฟโลฟ ดี.เอส., คาสุมยาน เอ.โอ. 2533. ฐานประสาทสัมผัสของพฤติกรรมการให้อาหารปลา // ประเด็น. วิทยา ต. 30. ฉบับที่. 5. หน้า 720-732.
85. พาฟโลฟ ดี.เอส., คาสุมยาน เอ.โอ. 2541. โครงสร้างพฤติกรรมการให้อาหารปลา // ประเด็น. วิทยา ต.38 ลำดับที่ 1 ป.123-136.
86. Pashchenko N.I. , Kasumyan A.O. พ.ศ. 2527 กระบวนการเสื่อมและการฟื้นฟูในเยื่อบุดมกลิ่นของปลาคาร์พหญ้า Ctenopharyngodon idella (Val.) (Cyprinidae) หลังจากการกระทำของผงซักฟอก Triton-X-100 กับมัน // ปัญหา วิทยา ต. 24. ประเด็น. 1. หน้า 128-137.
87. เพฟซเนอร์ พี.เอ. พ.ศ. 2521 การศึกษาด้วยกล้องจุลทรรศน์อิเล็กตรอนของต่อมรับรสของ Cytology ของปลาไหลยุโรป Anguilla anguilla II ต. 20 ลำดับ 10 หน้า 1112-1118
88. เพฟซเนอร์ พี.เอ. พ.ศ. 2523 ลักษณะวิวัฒนาการบางประการของการจัดระเบียบอวัยวะรับรสของปลา // ระบบประสาทสัมผัส กลิ่นและรสชาติ เจ.: วิทยาศาสตร์. หน้า 82-93.
89. เพฟซเนอร์ พี.เอ. 1981ก. โครงสร้างโครงสร้างของตัวรับรสในปลากระดูกพรุน I. ปลาสเตอร์เจียนที่โตเต็มวัย // เซลล์วิทยา ต. 23 ลำดับ 7 หน้า 760-766
90. เพฟซเนอร์ พี.เอ. 25259 การจัดระเบียบโครงสร้างของตัวรับรสในปลากระดูกพรุน I. ตัวอ่อนเปลี่ยนไปกินอาหารแบบแอคทีฟ // Cytology. ต. 23 ลำดับ 8 หน้า 867-873
91. เพฟซเนอร์ พี.เอ. 2528. โครงสร้างการจัดโครงสร้างของตัวรับรสในปลากระดูกพรุน. สาม. ตัวอ่อนในช่วงให้นมไข่แดง // Cytology. ต. 27 ลำดับ 11 หน้า 1240-1246
92. พอดดับนี เอ.จี. พ.ศ. 2509 ในการตอบสนองแบบปรับตัวของประชากรแมลงสาบต่อการเปลี่ยนแปลงสภาพที่อยู่อาศัย // Tr. สถาบันไบโอล น้ำภายใน สถาบันวิทยาศาสตร์แห่งสหภาพโซเวียต ฉบับที่ 10 (13)
93. ปราฟดิน ไอ.เอฟ. 2509. คู่มือการศึกษาปลา. อ.: จาก-ใน " อุตสาหกรรมอาหาร" 375 หน้า
94. ปุชคอฟ เอ็น.วี. 2497 สรีรวิทยาของปลา // Pishchepromizdat. ม. 371 น.
95. โรดิโอโนวา แอล.เอ. พ.ศ. 2512 การให้อาหารแมลงสาบในอ่างเก็บน้ำ Kama // บันทึกทางวิทยาศาสตร์ของมหาวิทยาลัยระดับการใช้งาน “ วัสดุของการประชุมทางวิทยาศาสตร์และการผลิตเกี่ยวกับสถานะของฐานวัตถุดิบของอ่างเก็บน้ำของภูมิภาคระดับการใช้งานและการใช้ประโยชน์ประมง” เพอร์เมียน
96. คู่มือเทคโนโลยีชีวภาพในการเพาะพันธุ์และเลี้ยงปลากินพืชเป็นอาหารตะวันออกไกล 2000. เอ็ด. วิโนกราโดวา วี.เค. VNIIPRH, 211 หน้า
97. Spanovskaya V.D. , Grigorash V.A. พ.ศ. 2504 จังหวะการให้อาหารไซปรินิดส์ทุกวัน // Vopr. นักวิทยาวิทยา ท.1 ฉบับ. 2 (19) หน้า 297-306.
98. สเตปาโนวา ฮ.เอ. พ.ศ. 2496 การใช้ประมงในอ่างเก็บน้ำ Katta-Kurgan // Izv. สถาบันวิทยาศาสตร์แห่ง UzSSR ฉบับที่ 5.
99. สทิจ ที.เอส. พ.ศ. 2516 การศึกษาทดลองการให้อาหารปลาคาร์พหญ้าวัยอ่อน // Kazakh Academy of Sciences สสส. นิเวศวิทยาของไฮโดรไบโอออนต์ในแหล่งน้ำของคาซัคสถาน อัลมา-อาตา. 136-142.
100. ซูตอฟ เอส.บี. 2482. สู่ความรู้เรื่องผลผลิตปลาในอ่างเก็บน้ำ. ข้อความที่ 8 ความสำคัญของตะกอนในการใช้อาหารธรรมชาติของปลา // Tr. ลิมนอล. ศิลปะ. ในโกสินทร์. ฉบับที่ 22.หน้า 241-249.
101. เฟโดรอฟ เอ.บี. พ.ศ. 2503 Ichthyofauna ของลุ่มน้ำ Don ในภูมิภาค Voronezh // ปลาและการประมงของภูมิภาค Voronezh (ผลงานของคณะสำรวจประมงบูรณาการปี 2496-2500) โวโรเนซ: จากมหาวิทยาลัยโวโรเนซ หน้า 149-249.
102. Flerova G.I., Gdovsky P.A. พ.ศ. 2519 ความเร็วของการแพร่กระจายของการกระตุ้นไปตามเส้นใยของเส้นประสาทรับกลิ่นและการตอบสนองที่เกิดขึ้นของสมองดมกลิ่นของปลาภายใต้สภาวะของอุณหภูมิที่เปลี่ยนแปลง // คำถามของ Ichthyology ต. 16. ประเด็น 1 (96) หน้า 119-125.
103. โฟคิน่า อี.เอส., กัสสุมยาน เอ.โอ. พ.ศ. 2546 การเปรียบเทียบความชอบด้านรสชาติในรุ่นต่างๆ ของประชากรของ Pungitius pungitius (Gasterosteiformes) ที่มีหนามเก้าหนาม // รายงานของ Academy of Sciences ต.389. ลำดับที่ 4. ป.570-573.
104. K.P. Fortunatova, Popova O.A. พ.ศ. 2516 ความสัมพันธ์ทางโภชนาการและอาหารของปลานักล่าในสามเหลี่ยมปากแม่น้ำโวลก้า อ.: วิทยาศาสตร์. 298 หน้า
105. Harborne D. 1985. ชีวเคมีสิ่งแวดล้อมเบื้องต้น. อ.: มีร์.289 น.
106. คาริโตโนวา เอ็น.เอช. 2506. ความสัมพันธ์ทางโภชนาการและอาหารของปลาคาร์พและปลาคาร์พสีเงินในบ่อ // Ukr. สถาบันวิจัยปลา. ครัวเรือน. "การส่งเสริม ผลผลิตปลาบ่อน้ำ” ทางวิทยาศาสตร์ ต. ต.XV. หน้า 7-25.
107. ไฮแอตต์ เค.ดี. 2526. กลยุทธ์โภชนาการ // พลังงานชีวภาพและการเจริญเติบโตของปลา. (เอ็ด. ดับบลิว. ฮวาเร และคณะ). อ.: อุตสาหกรรมเบา. หน้า 70-112.
108. เคเรมิโซวา เค.เอ. พ.ศ. 2501 โภชนาการของปลาไวท์ฟิชและปลาคาร์พสีเงินในแหล่งที่อยู่อาศัยใหม่ // Tr. สถาบันอุทกชีววิทยาเบลารุส ต.11.
109. ชเชอร์บีน่า จี.ค. พ.ศ. 2530 เรื่องการให้อาหารเทนช์บนโคลนสีเทาของทะเลสาบ Vishnetynetsky // ข้อมูล จดหมายข่าว " ชีววิทยาของน่านน้ำภายในประเทศ" ลำดับที่ 75. หน้า 43-47.
110. Shaposhnikova G.Kh. พ.ศ. 2507 ชีววิทยาและการแพร่กระจายของปลาในแม่น้ำประเภทอูราล อ.: วิทยาศาสตร์. 349 น.
111. ชิโวเคเน่ Y.S. 2532. การย่อยแบบ Symbiont ในสิ่งมีชีวิตในน้ำและแมลง. วิลนีอุส: ม็อกสลาส. 223 น.
112. โชริกิน เอ.เอ. 2495. ความสัมพันธ์ทางโภชนาการและอาหารของปลาในทะเลแคสเปียน. อ.: Pishchepromizdat. 267 น.
113. Aburto-Oropeza O., Sala E., Sanchez-Ortiz S. 2000. พฤติกรรมการให้อาหาร, การใช้แหล่งที่อยู่อาศัย, และความอุดมสมบูรณ์ของปลาเทวดา Holacanthus passer (Pomacanthidae) ในทะเลคอร์เตสตอนใต้ // ชีววิทยาสิ่งแวดล้อมของปลา. ว. 57. หน้า 435-442.
114. Adamek Z., Fasaic K., Debeljak L. 1990. ขีดจำกัดอุณหภูมิที่ต่ำกว่าของการบริโภคอาหารจากพืชในปลาคาร์พหญ้าอ่อน (Ctenopharyngodon idella Val.) // Ichthyologia หมายเลข 22. ป.1-8.
115. Adams M.A., Johnsen P.B., Hong-Qi Z. 1988. การเพิ่มประสิทธิภาพทางเคมีในการให้อาหารสำหรับปลาที่กินพืชเป็นอาหาร Tilapia zillii II การเพาะเลี้ยงสัตว์น้ำ ว. 72. หน้า 95-107.
116. Adron J.W., Mackie A.M. 2521 การศึกษาลักษณะทางเคมีของการให้อาหารสารกระตุ้นสำหรับเรนโบว์เทราต์ Salmo gairdneri Richardson // วารสารชีววิทยาปลา ว. 12. หน้า 303-310.
117. Appelbaum S. 1980. Versoche zur Geschmacksperzeption einiger imlarvalen undadulten Stadium//Arch. ฟิชเชอไรวิส. บด.31.เลขที่ 2 ป.105-114.
118. Atema J. 1971. โครงสร้างและหน้าที่ของความรู้สึกรับรสในปลาดุก (Ictalurus natalis) // สมอง, พฤติกรรมและวิวัฒนาการ. โวลต์ 4. หน้า 273-294.
119. Atema J. 1980. ความรู้สึกทางเคมี สัญญาณทางเคมี และพฤติกรรมการให้อาหารในปลา // พฤติกรรมของปลาและการใช้ในการจับและเพาะเลี้ยงปลา มะนิลา. ป.57-101.
120. เบิร์ด อาร์.ซี. 2508 ผลกระทบทางนิเวศวิทยาของพฤติกรรมของปลาบู่ Dimorphic ทางเพศ Microgobius gulosus (Girard) // Publ. สถาบัน วิทยาศาสตร์ทางทะเล เท็กซัส V.10.ป. 1-8.
121. บาร์โลว์ แอล.เอ., นอร์ธคัตต์ อาร์.จี. พ.ศ. 2538 ต้นกำเนิดของตัวอ่อนของปุ่มรับรสสัตว์ครึ่งบกครึ่งน้ำ // ชีววิทยาพัฒนาการ. ว. 169. หน้า 273-285.
122. บาร์ดาค เจ.อี., วินน์ เอช.อี., เมนเซล ดี.ดับเบิลยู. พ.ศ. 2502 ขั้วประสาทสัมผัสในการให้อาหารของนักล่ารีฟกลางคืน Gymnothorax moringa และ G.vicinus // Copeia ลำดับที่ 2 ป.133-139.
123. บาร์ดัค เจ.อี., ท็อดด์ เจ.เอช., คริกเมอร์ อาร์.เค. 2510. การจัดวางตามรสชาติของปลาในสกุล Ictalurus II Science. ลำดับที่ 155 ป.276-1278.
124. ไบด์เลอร์ แอล.เอ็ม. 2510 ประจุลบมีอิทธิพลต่อการตอบสนองของตัวรับรส // การดมกลิ่นและรสชาติ II (เอ็ด. ที. ฮายาชิ) อ็อกซ์ฟอร์ด: Pergamon Press. หน้า 509-535.
125. Brown S.B., Evans R.E., Thompson B.E., Hara T.J. 2525. การรับเคมีบำบัดและมลพิษทางน้ำ // การรับสารเคมีในปลา (Ed. T.J. Hara) บริษัท เอลส์เวียร์ ไซแอนติฟิค พับลิชชิ่ง อัมสเตอร์ดัม ป.363-393.
126. ไบรอัน เจ.อี., ลาร์คิน พี.เอ. 2515 ความเชี่ยวชาญด้านอาหารโดยปลาเทราท์แต่ละตัว // วารสารคณะกรรมการวิจัยการประมงแห่งแคนาดา ว. 29. ป. 1615-1624.
127. คัลแลน ดับเบิลยู.ที., แซนเดอร์สัน เอส.แอล. 2546. กลไกการให้อาหารในปลาคาร์พ: การกรองแบบ crossflow, การยื่นออกของเพดานปากและการกลับตัวของการไหล // J. Exp. ไบโอล โวลต์ 206. หน้า 883-892.
128. Caprio J. 1975. ความไวสูงของตัวรับรสปลาดุกต่อกรดอะมิโน // J. Comp. ไบโอเคม ฟิสิออล. V. 52 A.P. 217-251.
129. Caprio J. 1978. การดมกลิ่นและรสชาติในปลาดุกแชนเนล: การศึกษาทางไฟฟ้าสรีรวิทยาของการตอบสนองต่อกรดอะมิโนและอนุพันธ์ // วารสารสรีรวิทยาเปรียบเทียบ. ว. 123. หน้า 357-371.
130. Dabrowski K., Rusiecki M. 1983. เนื้อหาของกรดอะมิโนทั้งหมดและอิสระในอาหารแพลงก์ตอนสัตว์ของตัวอ่อนปลา // การเพาะเลี้ยงสัตว์น้ำ. ว.30. ข้อ 1-4. ป.31-42.
131. De la Noue J., Choubert G. 1985. การย่อยได้ชัดเจนของชีวมวลที่ไม่มีกระดูกสันหลังด้วยเรนโบว์เทราต์ // การเพาะเลี้ยงสัตว์น้ำ. ว. 50. หน้า 103-112.
132. เดวิสินา จี.วี. 2546. เรื่องปฏิสัมพันธ์ของระบบเคมีเซ็นเซอร์ในปลา // J. of Ichthyology, V.43. อุปทาน 2. “พฤติกรรม การแพร่กระจายและการอพยพของปลา”. ป.214-227.
133. โดวิ่ง K.B. 2529. คุณสมบัติเชิงหน้าที่ของระบบรับกลิ่นของปลา. // ความก้าวหน้าทางสรีรวิทยาทางประสาทสัมผัส. โวลต์ 6. เบอร์ลิน ป.39-104.
134. Doving K.B., Seslet R., Tommesen G. 1980. ความไวในการดมกลิ่นต่อกรดน้ำดี ปลา insalmonid // Acta Physiol. สแกน ว. 108. หน้า 123-131.
135. Dubois-Dauphin M., Doving K.B., Holley A. 1980. ความสัมพันธ์ทางภูมิประเทศระหว่างป่องดมกลิ่นกับทางเดินดมกลิ่นในเทนช์ (เกลื้อน tinea L) // ความรู้สึกทางเคมี. ว.5.ข้อ 2.ป.159-169.
136. ดัสซอลต์ จี.วี., เครเมอร์ ดี.แอล. พ.ศ. 2524 พฤติกรรมการกินอาหารและการให้อาหารของปลาหางนกยูง Poecilia reticulata (Pisces: Poeciliidae) // Canadian Journal of Zoology. ว. 59. หน้า 684-701.
137. Farr J.A., Herrnkind W.F. พ.ศ. 2517 การวิเคราะห์เชิงปริมาณของปฏิสัมพันธ์ทางสังคมของปลาหางนกยูง Poecilia reticulate (Pisces: Poeciliidae) เป็นหน้าที่ของความหนาแน่นของประชากร // พฤติกรรมของสัตว์ ลำดับที่ 22 น. 582-591.
138. นิ้ว T.E. 2519. วิถีการกินของปลาดุกหัวกระทิง. ตอนที่ 1. การเชื่อมต่อของปมประสาทส่วนหน้า // วารสารประสาทวิทยาเปรียบเทียบ. ว. 165. หน้า 513-526.
139. Finger T.E., Morita Y. 2528. ระบบรับรสสองระบบ: นิวเคลียสรับรสของใบหน้าและช่องคลอดมีการเชื่อมต่อก้านสมองที่แตกต่างกัน // วิทยาศาสตร์ ว. 227. หน้า 776778.
140. Finger T.E., Drake S.K., Kotrschal K., Womble M., Dockstader K.C. 1991. การเจริญเติบโตหลังตัวอ่อนของระบบรับรสส่วนปลายในปลาดุกช่อง, Ictalurus punctatus II Journal of Comparative Neurology. ลำดับที่ 314 ป.55-66.
141. แฟรงก์ M.E., Hettinger T.P., Mott A.E. พ.ศ. 2535 ความรู้สึกในการรับรส: ชีววิทยาทางระบบประสาท อายุ และผลกระทบของยา // บทวิจารณ์เชิงวิพากษ์ทางชีววิทยาช่องปากและการแพทย์ V. 3. หมายเลข 4. หน้า 371-393.
142. Frankiewicz P, Zalewski M., Biro P, Tatrai I, Przybylki M. 1991. อาหารปลาจากลำธารทางตอนเหนือของพื้นที่กักเก็บน้ำของทะเลสาบ Balaton (ฮังการี) // Acta Hydrobiol, V. 33 . หมายเลข 1-2. ป.149-160.
143. Gerhart D.J., Bondura M.E., Commito J.A. พ.ศ. 2534 การยับยั้งการให้อาหารปลาซันฟิชโดยสเตียรอยด์ป้องกันจากแมลงเต่าทอง: ความสัมพันธ์ระหว่างโครงสร้างกับกิจกรรม // วารสารนิเวศวิทยาเคมี. ว. 17. ป. 1363-1370.
144. โกห์ อี ทามูระ ต. 1980a. ผลของกรดอะมิโนต่อพฤติกรรมการให้อาหารในลมหายใจทะเลแดง // ชีวเคมีและสรีรวิทยาเปรียบเทียบ 66ซี ป.225-229.
145. โกห์ อี ทามูระ ต. 1980b. การตอบสนองทางจมูกและการรับรสต่อกรดอะมิโนในลมหายใจทะเลแดงและปลากระบอก 2 ชนิด // ชีวเคมีและสรีรวิทยาเปรียบเทียบ 66ซี ป.217-224.
146. Gomahr A, Palzenberger M, Kotrschal K. 1992. ความหนาแน่นและการกระจายของต่อมรับรสภายนอกในไซปรินิดส์ // สภาพแวดล้อม ไบโอล ปลา. ว. 33. ลำดับที่ 12. หน้า 125-134.
147. ฮัลเพอร์น บี.พี. พ.ศ. 2529 ข้อ จำกัด ที่กำหนดเกี่ยวกับสรีรวิทยาของรสชาติโดยข้อมูลเวลาปฏิกิริยาของรสชาติของมนุษย์ // Neurosci ประพฤติตน ความละเอียด ว. 10. หน้า 135-151.
148. Hara T.J., Sveinsson T., Evans R.E., Klaprat D.A. พ.ศ. 2536 ลักษณะทางสัณฐานวิทยาและการทำงานของอวัยวะรับกลิ่นและการรับรสของ Salvelinus สามสายพันธุ์ // Can. เจ ซูล. V. 71. ลำดับ 2. หน้า 414-423.
149. Hara T.J., Carolsfeld J., Kitamura S. 1999. ความแปรปรวนของความรู้สึกรับรสในปลาแซลมอน โดยมีการอ้างอิงพิเศษถึงความแตกต่างของสายพันธุ์ในเรนโบว์เทราท์, Oncorhynchus mykiss // Can เจ.ฟิช. น้ำ. วิทยาศาสตร์ ว. 56. ป. 13-24.
150. ฮาร์ต P.J.B. , กิลล์ เอ.บี. 2535 ข้อจำกัดในการเลือกขนาดเหยื่อโดย Stickleback แบบสามเข็ม: ความต้องการพลังงาน ความจุ และความสมบูรณ์ของลำไส้ J. Fish Biol ว. 40. หน้า 205-218.
151. ไฮนส์โบรค แอล.ที.เอ็น., ครูเกอร์ เจ.จี. 2535. การให้อาหารและการเจริญเติบโตของปลาไหลแก้ว, Anguilla anguilla L. ผลของการให้อาหารสารกระตุ้นต่อการกินอาหาร การเผาผลาญพลังงาน และการเจริญเติบโต // การจัดการการเพาะเลี้ยงสัตว์น้ำและการประมง ลำดับที่ 23 น. 327336
152. Hellstrom T., Doving K.B. 2529. การรับเคมีบำบัดของทอโรโคเลทในปลาคอดที่ผ่าตัดด้วย anosmic และเสแสร้ง, Gadus morhua // การทำงานของสมองเชิงพฤติกรรม. ว.21.ป.155-162.
153. เฮอร์ริค ซี.เจ. พ.ศ. 2444 เส้นประสาทสมองและอวัยวะรับสัมผัสทางผิวหนังของปลาไซลูริดในอเมริกาเหนือ // วารสารประสาทวิทยาและสรีรวิทยาเปรียบเทียบ ว. 11. หน้า 177-249.
154. Hidaka I. 1982. การกระตุ้นตัวรับรสชาติและพฤติกรรมการให้อาหารในปลาปักเป้า // Chemoreception ในปลา (เอ็ด. ที.เจ. ฮารา). บริษัท เอลส์เวียร์ ไซแอนติฟิค พับลิชชิ่ง อัมสเตอร์ดัม ป.243-257.
155. Hidaka I., Ishida Y. 1985. การตอบสนองของ Gustatoiy ในแถลงการณ์ Shimaisaki (ปลาเสือ) Therapon oxyrhynchus II ของสมาคมประมงวิทยาศาสตร์แห่งญี่ปุ่น ว. 51. หน้า 387-391.
156. Hidaka I., Ohsugi T., Kubomatsu T. 1978. การกระตุ้นตัวรับรสชาติและพฤติกรรมการกินอาหารในปลาปักเป้า Fugu pardalis ส่วนที่ 1 ผลของสารเคมีชนิดเดียว // ความรู้สึกและรสชาติทางเคมี ลำดับที่ 3 ป.341-354.
157. Holm J.C., Walther B. 1988. กรดอะมิโนอิสระในแพลงก์ตอนสัตว์น้ำจืดและอาหารแห้ง: ความสำคัญที่เป็นไปได้สำหรับการให้อาหารครั้งแรกในปลาแซลมอนแอตแลนติก (Salmo salar) // Aquakulture. ลำดับที่ 71. ป.341-354.
158. Horppila J. 1994. อาหารและการเจริญเติบโตของแมลงสาบ (Rutilus rutilus (L.)) ในทะเลสาบ Vesijarvi และการเปลี่ยนแปลงที่เป็นไปได้ในกระบวนการจัดการทางชีวภาพ // Hydrobiologia ว. 294. หน้า 35-41.
159. Horppila J. 1999. การเปลี่ยนแปลงองค์ประกอบอาหารของ Cyprinid ที่กินไม่ได้ทุกชนิดซึ่งเป็นแหล่งที่มาของข้อผิดพลาดที่เป็นไปได้ในการประมาณการบริโภคอาหาร // Hydrobiologia ว. 294. หน้า 35-41.
160. Horppila J., Ruuhijarvi J., Rask M., Karppinen C., Nyberg K., Olin M. 2000. การเปลี่ยนแปลงตามฤดูกาลในอาหารและความอุดมสมบูรณ์ของคอนและแมลงสาบในบริเวณชายฝั่งและทะเลของทะเลสาบขนาดใหญ่ / / วารสารชีววิทยาปลา. ว. 56. ป. 51-72.
161. Jakubowski M. 1983. รายละเอียดใหม่ของโครงสร้างพิเศษ (TEM, SEM) ของต่อมรับรสในปลา // Zeitschrift fur Mikroskopisch-Anatomische Forschung ว. 97. หน้า 849-862.
162. Jakubowski M. , Whitear M. 1990. สัณฐานวิทยาเปรียบเทียบและเซลล์วิทยาของต่อมรับรสใน teleosts // Z. mikrosk.-anat ฟอร์ช. ว. 104. ลำดับ 4. หน้า 529-560.
163. โจนส์ เค.เอ. 2532. ความอร่อยของกรดอะมิโนและสารประกอบที่เกี่ยวข้องกับเรนโบว์เทราท์, Salmo gairdneri Richardson // J. Fish Biol. ว. 34. ที่ ลพ. 149-160.
164. คาคุ ต., สึมาการิ เอ็ม., คิโยฮาระ เอส., ยามาชิตะ 2523. การตอบสนองอย่างน่ารับประทานในสร้อย Pseudorasbora parva I I สรีรวิทยาและพฤติกรรม. ว.25.ป.99-105.
165. คัมสตรา เอ., ไฮนส์บรูค แอล.ที.เอ็น. 2534. ผลของตัวดึงดูดต่อการเริ่มให้อาหารปลาไหลแก้ว, Anguilla anguilla L. // การจัดการการเพาะเลี้ยงสัตว์น้ำและการประมง. ลำดับที่ 22. ป.47-56.
166. Kanwal J.S., Caprio J. 1983. การตรวจสอบทางสรีรวิทยาของระบบการรับรสของช่องปาก (IX-X) ในช่องปลาดุก Ictalurus punctatus II J. Comp. ฟิสิออล. ก.V. 150. หน้า 345-357.
167. Kanwal J.S. , Caprio J. 1988. รสชาติที่ทับซ้อนกันและแผนที่สัมผัสของ oropharynx ในกลีบช่องคลอดของปลาดุกช่อง Ictalurus punctatus // J. Neurobiol ว. 19. ลำดับ 3. ป. 211-222.
168. กันวาล เจ.เอส., ฟิงเกอร์ ที.อี. พ.ศ. 2535 การเป็นตัวแทนและการฉายภาพส่วนกลางของระบบรับรส // Fish Chemoreception (Ed. T.J.Hara) แชปแมนและฮอลล์ ลอนดอน. ป.79-102.
169. Kapoor B.G., Evans H.E., Pevzner R.A. พ.ศ. 2518 ระบบการรับรสของปลา // พัฒนาชีววิทยาทางทะเล ว.13.ป.53-108.
170. Kasumyan A.O., 2542. การดมกลิ่นและการรับรสในพฤติกรรมของปลาสเตอร์เจียน. // เจ Appl. อิคธิออล. ว. 15. หน้า 228-232.
171. เกษมยาน อ.โอ. 2545. พฤติกรรมการค้นหาอาหารปลาสเตอร์เจียนที่เกิดจากสิ่งเร้าทางเคมี: กลไกทางประสาทสัมผัสที่เชื่อถือได้ // J. Appl. อิคธิออล. ว. 18. ป. 685-690.
172. เกษมยาน อ.โอ. 2547. ระบบรับกลิ่นในปลา: โครงสร้าง หน้าที่ และบทบาทในพฤติกรรม//เจ. อิคธิออล. ว. 44. อุปทาน. 2. หน้า 180-223.
173. Kasumyan A.O., Doving K.B. 2546. ความชอบด้านรสชาติของปลา // ปลาและการประมง. โวลต์ 4. หน้า 289-347.
174. Kasumyan A.O., Nikolaeva E.V. 2545. การวิเคราะห์เปรียบเทียบความชอบด้านรสชาติของปลาที่มีระบบนิเวศและการให้อาหารต่างกัน // Journal of Ichthyology. ฉบับที่ 41. อุปทาน 2. หน้า 203-214.
175. Kasumyan A.O., Sidorov S.S., 1995. ความอร่อยของกรดอะมิโนอิสระและสารรสชาติคลาสสิกใน Frolich Char, Salvelinus alpinus erhythrinus (Georgi) // นอร์ดิก เจ. เฟรชว. ความละเอียด ลำดับที่ 71 หน้า 320-323
176. Kasumyan A.O. , Sidorov S.S. 2545 ความแปรปรวนของความชอบส่วนบุคคลในปลาสร้อย Phoxinus phoxinus II Journal of Ichthyology ฉบับที่ 42. อาหารเสริม 2. หน้า 241-254.
177. Kasumyan A.O., Marusov E.A., Sidorov S.S., 2003. พฤติกรรมการให้อาหารของ ruffe Gymnocephalus cernuus ที่ถูกกระตุ้นโดยสารกระตุ้นการดมกลิ่นและการรับรส วารสารวิทยา. ว. 43. อุปทาน. 2. หน้า 247-254.
178. Kennedy M., Fitzmaurice P. 1970. ชีววิทยาของเทนช์, เกลื้อน tinea (L.) ในน่านน้ำไอริช // Proc. รอยัลไอริชอคาด ว. 69. ป.31-82.
179. Kiyohara S., Yamashita S., Harada S. 1981. ความไวสูงของตัวรับรส minnow ต่อกรดอะมิโน // Physiol, และ Behav. V. 26. ลำดับ 6. หน้า 1103-1108.
180. คิโยฮาระ เอส., ชิราทานิ ต., ยามาชิตะ เอส.1985. การกระจายบริเวณรอบนอกและส่วนกลางของกิ่งก้านหลักของเส้นประสาทรับรสใบหน้าในปลาคาร์พ // Brain Res. ว. 325. หน้า 57-69.
181. Klaprat D.A., Evans R.E., Hara T.J. 2535. สารปนเปื้อนในสิ่งแวดล้อมและการรับเคมีบำบัดในปลา // ใน: การรับเคมีบำบัดของปลา (ed. T.J.Hara) แชปแมนและฮอลล์ ลอนดอน. ป.321-341.
182. Kleerekoper H. 1969. การดมกลิ่นในปลา. บลูมิงตัน มหาวิทยาลัยอินเดียนา กด.
183. Konishi J. Zotterman Y. 1961. การทำงานของรสชาติในปลาคาร์พ: การศึกษาทางไฟฟ้าสรีรวิทยาเกี่ยวกับเส้นใยรับรส // Acta Physiologica Scandinavica. ว. 52. หน้า 150-161.
184. Kotrschal K., Peters R.C., Doving K.B. 2539. การตอบสนองทางเคมีและประสาทสัมผัสจากครีบหลังด้านหน้าของหินโยก, Gaidropsarus vulgaris (Gadidae, Teleostei) // Prim.Sensory Neuron V. 1. หมายเลข 4. หน้า 297-309.
185. แม็กกี้ A.M. 2525. การจำแนกสารกระตุ้นการกินอาหาร // การรับเคมีบำบัดในปลา. (เอ็ด. ที.เจ. ฮารา). เอเลเซเวียร์ ไซแอนติฟิค บมจ. คอมพ์ อัมสเตอร์ดัม ป.275-291.
186. แม็กกี้ เอ.เอ็ม., เอดรอน เจ.ดับบลิว. 2521 การจำแนก inosine และ inosine-5"-monophosphate เป็นตัวกระตุ้นการให้อาหารสำหรับ turbot, Scophthalmus maximus // ชีวเคมีและสรีรวิทยาเปรียบเทียบ 60 A. P. 79-88
187. Mackie A.M., Mitchell A.I. 2526. การศึกษาลักษณะทางเคมีของสารกระตุ้นการให้อาหารสำหรับปลาไหลยุโรปรุ่นเยาว์, แองกวิลลา แองกวิลลา (L) // วารสารชีววิทยาปลา. ว. 22. ป 425-430.
189. Marui T., Caprio J. 1992. Teleost gustation // ใน: Fish Chemoreception (Ed. T.J.Hara) แชปแมนและฮอลล์ ลอนดอน. ป.171-198.
190. มารุย ที., อีแวนส์ อาร์., ซีลินสกี้ บี., ฮารา ที.เจ. 2526. การตอบสนองอย่างน่ารับประทานของปลาเรนโบว์เทราต์ (Salmo gairdneri) ต่อกรดอะมิโนและอนุพันธ์ // เจคอมพ์. ฟิสิออล. ว. 153เอ. ป.423-433.
191. Mearns K.J., Ellingsen O.F., Doving K.B., Helmer S. 1987. พฤติกรรมการให้อาหารในเรนโบว์เทราต์สำหรับผู้ใหญ่และปลาแซลมอนแอตแลนติกพาร์ ระบุโดยเศษส่วนทางเคมีและส่วนผสมของสารประกอบที่ระบุในสารสกัดจากกุ้ง // การเพาะเลี้ยงสัตว์น้ำ ลำดับที่ 64. ป.47-63.
192. มิสเตรตต้า ซี.เอ็ม. 2534. ชีววิทยาพัฒนาการของระบบรับรส // ใน: กลิ่นและรสชาติในสุขภาพและโรค. นิวยอร์ก: Raven Press ป.35-64.
193. Nikolaeva E.V., Kasumyan A.O. 2000. การวิเคราะห์เปรียบเทียบความชอบด้านรสชาติและการตอบสนองเชิงพฤติกรรมต่อสิ่งเร้าทางลมในตัวเมียและตัวผู้ของปลาหางนกยูง Poecilia reticulata II Journal of Ichthyology ว. 40. หน้า 479-484.
194. Ohsugi T., Hidaka I., Ikeda M. 1978. การกระตุ้นตัวรับรสชาติและพฤติกรรมการกินอาหารในปลาปักเป้า, Fugu pardallis. ส่วนที่ 2 ผลที่เกิดจากส่วนผสมของสารสกัดจากหอย // ความรู้สึกทางเคมีและรสชาติ ว. 3. ป. 355-368.
195. O'Maoileidigh N. , Bracken J.J. 2532. ชีววิทยาของเทนช์, เกลื้อน tinea (L. ) ในทะเลสาบไอริช // การจัดการการเพาะเลี้ยงสัตว์น้ำและการประมง V. 20. ลำดับ 2. หน้า 199-209
196. ออสเซ่ เจ.ดับเบิลยู.เอ็ม., ซิบบิง เอฟ.เอ., ฟาน เดน บูการ์ต เจ.จี.เอ็ม. 2540. การจัดการอาหารในช่องปากของปลาคาร์พและ pther cyprinids: การปรับตัวและข้อ จำกัด // Acta Physiol. สแกน ว. 161. อุปทาน. 638. หน้า 47-57.
197. Perkar C., Krupauer V. 1968. ความสัมพันธ์ทางอาหารระหว่างปลาคาร์พอายุ 2 ปีกับเทนช์ในสต็อกหลายชนิดผสม // Prace VURN Vodn., N8, P.29-54.
198. รามิเรซ ไอ. สปร็อตต์ อาร์.แอล. พ.ศ. 2521 กลไกทางพันธุกรรมของการดื่มและการให้อาหาร // Neurosci. ไบโอบีฮาฟ สาธุคุณ V.2.เลขที่ 1. ป.15-26.
199. Reid M., Hammersley R. 1996. ผลของเครื่องดื่มอัดลมต่อพฤติกรรมการกินในช่วงเจ็ดวัน // Proc. นูทริ สังคมสงเคราะห์ ว.55.ฉบับที่. 3. หน้า 251.
200. Reiter R., Lukowicz M.v., Arnold R., le Deit H., Aquaron R., Schmidter A., Kuznik M., Burkard S., Rannz D., Rambeck W.A. 2545 Algen im Fischfutter -eine Möglichkeit der Jochanreicherung im Süßwasserfisch // Fischer und Teichwirt ว. 53. ลำดับที่ 6. ป.211-212
201. ริงเลอร์ เอ็น.เอช. พ.ศ. 2528 ความแปรผันส่วนบุคคลและเวลาในการเปลี่ยนเหยื่อโดยปลาเทราต์สีน้ำตาล Salmo trutta // Copea โวลต์ 4. หน้า 918-926.
202. Reutter K. 1971. Die Geschmacksknospen des Zwergwelses Amiurus nebulosus, Morphologische und histochemixche Unterschungen // Z. mikr. อานนท์. บด. 120. ส. 280-308.
203. Reutter K. 1986. ตัวรับเคมี // ใน: ชีววิทยาของจำนวนเต็ม. V.II. (Ed. J. Bereiter-Hahn, A. G. Matoltsy และ K. S. Richards) เบอร์ลิน สปริงเกอร์. ป.586-604.
204. Reutter K. 1992. โครงสร้างของอวัยวะรับรสที่แสดงโดยปลาไซลูรอยด์ Plotosus lineatus (Thunberg) // Fish Chemoreception (Ed. T.J.Hara) แชปแมนและฮอลล์ ลอนดอน. ป.60-78.
205. Reutter K., Witt M. 1993. สัณฐานวิทยาของอวัยวะรับรสของสัตว์มีกระดูกสันหลังและเส้นประสาท // ใน: กลไกของการถ่ายโอนรสชาติ (Ed. S.A. Simon และ S.D. Roper) ซีอาร์ซี เพรส. โบคา ราตัน. ป.29-82.
206. Sakashita H. 1992. พฟิสซึ่มทางเพศและนิสัยการกินอาหารของปลา clingfish, Diademichthys lineatus และการพึ่งพาอาศัยเม่นทะเลที่เป็นเจ้าภาพ // สภาพแวดล้อม ไบโอล ปลา. ว. 34. ป. 95-101.
207. ชูลท์ บ., บาคุส จี.เจ. พ.ศ. 2535 การยับยั้งการปล้นสะดมในฟองน้ำทะเล: ห้องปฏิบัติการกับการศึกษาภาคสนาม // บูล วิทยาศาสตร์ทางทะเล ว. 50. ลำดับที่ 1. หน้า 205-211.
208. เซลเซต อาร์. โดวิ่ง เค.บี. 2523. พฤติกรรมของถ่านอะนาโดรมัสที่โตเต็มวัย (Salmo alpinus L.) ต่อกลิ่นที่เกิดจากการลอกคราบของประชากรพวกมันเอง // Acta Physiol. สแกน ว. 108. หน้า 113-122.
209. พี่น้องเอฟ.เอ. 2531. ความเชี่ยวชาญและข้อจำกัดในการใช้ทรัพยากรอาหารโดยปลาคาร์พ Cyprinus carpio: การศึกษาการแปรรูปอาหารในช่องปาก // ชีววิทยาสิ่งแวดล้อมของปลา. ลำดับที่ 22. ป.161-178.
210. Sibbing F.A., Osse J.W.M., Terlouw A. 2529. การจัดการอาหารในปลาคาร์พ (Cyprinus carpio): รูปแบบการเคลื่อนไหว กลไกและข้อ จำกัด // J. Zool สังคมสงเคราะห์ ของลอนดอน ว. 210 (ก) ลำดับที่ 2 ป.161-203.
211. ซินแคลร์ เจ.ดี., กัมโปฟ-โพเลวอย เอ., สจ๊วร์ต อาร์., ลี ที.-เค. พ.ศ. 2535 รสนิยมในบรรทัดหนูที่เลือกสำหรับการบริโภคแอลกอฮอล์ต่ำและสูง // แอลกอฮอล์ ว. 9. ลำดับที่ 2. ป.155-160.
212. Sukop I., Adamek Z. 2538. ชีววิทยาอาหารของเทนช์อายุ 1, 2 และ 3 ปีในการเลี้ยงแบบผสมผสานกับปลาคาร์พและปลากินพืช // พล. โค้ง. ไฮโดรไบโอล ว.42. ข้อ 1-2. ป.9-18.
213. ซัทเทอร์ลิน A.M. พ.ศ. 2518 แรงดึงดูดทางเคมีของปลาทะเลบางชนิดในแหล่งที่อยู่อาศัยตามธรรมชาติ // วารสารคณะกรรมการวิจัยการประมงแห่งแคนาดา ลำดับที่ 32 หน้า 729-738
214. Sutterlin A.M., Sutterlin N. 1970. รสชาติที่ตอบสนองต่อปลาแซลมอน Arlantic (Salmo salar) parr // วารสารคณะกรรมการวิจัยการประมงแห่งแคนาดา. ลำดับที่ 27 น. 2470-2485
215. Takeda M., Takii K. 1992. รสชาติและโภชนาการในปลา: การประยุกต์กับการเพาะเลี้ยงสัตว์น้ำ // ใน: Fish Chemoreception (Ed. T.J. Hara) แชปแมนและฮอลล์ ลอนดอน. ป.271-287.
216. Takeda M., Takii K., Matsui K. 1984. การระบุสารกระตุ้นการให้อาหารสำหรับปลาไหลเด็กและเยาวชน // แถลงการณ์ของสมาคมประมงวิทยาศาสตร์แห่งญี่ปุ่น. ลำดับที่ 50 หน้า 1039-1043
217. Takii K., Shimeno S., Takeda M., Kamekawa S. 1986. ผลของการให้อาหารสารกระตุ้นในอาหารต่อกิจกรรมเอนไซม์ย่อยอาหารของปลาไหล // กระดานข่าวของสมาคมประมงวิทยาศาสตร์แห่งญี่ปุ่น. ลำดับที่ 52 ป.1449-1454.
218. Valentincic T., Caprio J. 1994. พฤติกรรมการให้อาหารกรดอะมิโนในสมบูรณ์และปลาดุกช่อง anosmic Ictalurus punctatus // Physiol. ประพฤติตน ว. 55. ลำดับที่ 5. หน้า 857-863.
219. แวน แดมม์ อาร์., เบาเวนส์ ดี., แวนเดอร์สทิเฮเลน ดี., เวอร์เฮเยน อาร์.เอฟ. 2533 การตอบสนองของจิ้งจก Lecerta vivípara ต่อสัญญาณเคมีนักล่า: ผลกระทบของอุณหภูมิ // พฤติกรรมสัตว์. ว. 40. หน้า 298-305.
220. เวเธอร์ลีย์ N.S. พ.ศ. 2530 อาหารและการเจริญเติบโตของ dace 0 กลุ่ม Leuciscus leuciscus (L. ) และแมลงสาบ Rutilus rutilus (L. ) ในแม่น้ำที่ราบลุ่ม // J. Fish Biol ว. 30. ป. 237-247.
221. เวเบอร์ อี.เอช. พ.ศ. 2370 (ค.ศ. 1827) Über das Geschmacksorgane des Karpfen und den Ursprung seiner Nerven เอกสารสำคัญสำหรับกายวิภาคศาสตร์และสรีรวิทยา ป.309-315.
222. Welsch IL, Storch V. 1969. Die Feinstruktur der Geschmacksknospen von Welsen Ciarias batrachus (L.) และ Kryptopterus bicirrhis (Cuvier et Valenciennes) // Zeitschrift für Zellforschung และ Mikroscopische Anatomie ว. 100. หน้า 552-559.
223. Witt M., Reutter K. 1990. การสาธิตด้วยกล้องจุลทรรศน์อิเล็กตรอนของบริเวณจับกับเลคตินในต่อมรับรสของปลาดุกยุโรป Silurus glanis (Teleostei) // ฮิสโตเคมี. ว.94 ป.617-628.
224. White A.W., Fukuhara O., Anraku M.1989. การตายของลูกปลาจากการกินสารพิษไดโนแฟลเจลเลต // กระแสน้ำแดง: ชีววิทยา วิทยาศาสตร์สิ่งแวดล้อม และพิษวิทยา โปรค อินเตอร์ที่ 1 อาการ กระแสน้ำสีแดง นิวยอร์ก. ป.395-398.
225. Whitear M. 1971 ความเชี่ยวชาญเฉพาะทางของเซลล์และการทำงานของประสาทสัมผัสในหนังกำพร้าของปลา // วารสารสัตววิทยา ลอนดอน. ว. 163. หน้า 237-264.
226. Whitear M. 1992. เซลล์เคมีบำบัดเดี่ยว // ใน: Fish Chemoreception (Ed. T.J. Hara) แชปแมนและฮอลล์ ลอนดอน. ป.103-125.
227. วูตตัน อาร์.เจ. 2541 นิเวศวิทยาของปลา Teleost // ใน: สำนักพิมพ์ทางวิชาการของ Kluwer ดอร์เดรชท์.
228. Wunder W. 1957. Die Sinnesorgane der Fische // Allgem ฟิชเชอร์ไรซ์ตุง. ว. 82. ป. 1-24.
229. Yamamoto T., Kawamura Y. 1981. เวลาตอบสนองของลมในผู้ใหญ่ของมนุษย์ // Physiol. ประพฤติตน ว. 26. หน้า 715-719.
230. Yoshii K., Kamo N., Kurihara K., Kabataki Y. 1979. การตอบสนองอย่างน่ารับประทานของตัวรับเพดานปากปลาไหลต่อกรดอะมิโนและกรดคาร์บอกซิลิก // วารสารสรีรวิทยาทั่วไป. ว. 74. หน้า 301-317.
231. Zuwala K., Jakubowski M. 1993. กล้องจุลทรรศน์แสงและอิเล็กตรอน (SEM, TEM) ของต่อมรับรสในเทนช์ Tinca tinea (ราศีมีน: Cyprinidae) II Acta Zoologica สตอกโฮล์ม V. 74. ลำดับ 4. หน้า 277-282.
โปรดทราบว่าข้อความทางวิทยาศาสตร์ที่นำเสนอข้างต้นถูกโพสต์เพื่อวัตถุประสงค์ในการให้ข้อมูลเท่านั้น และได้รับผ่านการจดจำข้อความวิทยานิพนธ์ต้นฉบับ (OCR) ดังนั้นอาจมีข้อผิดพลาดที่เกี่ยวข้องกับอัลกอริธึมการรู้จำที่ไม่สมบูรณ์
ไม่มีข้อผิดพลาดดังกล่าวในไฟล์ PDF ของวิทยานิพนธ์และบทคัดย่อที่เราจัดส่ง
แนวคิดเรื่องรสนิยมทางสุนทรีย์นั้นเกิดขึ้นในวัฒนธรรมยุโรปในยุคประวัติศาสตร์ที่ค่อนข้างช้าบนพื้นฐานของประสบการณ์ทางจิตวิญญาณที่เป็นรายบุคคลและกลายเป็นเงื่อนไขสำหรับความหลากหลายของเนื้อหาของคุณค่าทางจิตวิญญาณ รสนิยมทางสุนทรีย์หมายถึงความสามารถของบุคคลในการรับรู้และประเมินระดับความสมบูรณ์ทางสุนทรียภาพของโลกวัตถุประสงค์และปรากฏการณ์ทางจิตวิญญาณ ทั้งนี้ขึ้นอยู่กับความรู้สึกยินดีหรือไม่พอใจ รสนิยมทางสุนทรีย์สะท้อนให้เห็นตัวเองในการตัดสินคุณค่า เช่นเดียวกับในกิจกรรมที่สร้างสรรค์ทุกประเภท ตั้งแต่การแสดงออกถึงสไตล์ในชีวิตประจำวันในเสื้อผ้า วิถีชีวิต ที่เกี่ยวข้องกับคุณค่าทางสังคม โดยเฉพาะอย่างยิ่งทางศิลปะ คำจำกัดความคลาสสิกของรสนิยมทางสุนทรีย์มีอยู่ในผลงานของคานท์เรื่อง “มานุษยวิทยาในความสัมพันธ์เชิงปฏิบัติ” นักปรัชญาเขียนว่า “รสชาติคือความสามารถของความสามารถในการตัดสินเชิงสุนทรีย์ในการตัดสินใจเลือกที่มีความสำคัญสากล” ดังที่นักปรัชญาตั้งข้อสังเกต เรากำลังพูดถึงความสอดคล้องของการตัดสินของเรากับคุณสมบัติที่เป็นวัตถุประสงค์ของหัวข้อการตัดสิน ซึ่งเป็นกุญแจสำคัญสู่ความเป็นสากล ความเที่ยงธรรมของการตัดสินรสชาตินั้นพิสูจน์ได้จากการมีประสบการณ์ด้านสุนทรียศาสตร์ที่พัฒนาแล้วซึ่งสัมพันธ์กับโลก บนพื้นฐานนี้ นักปรัชญาชาวเยอรมัน I. Sulzer ได้พัฒนารสนิยมควบคู่ไปกับความสามารถทางปัญญา เช่น ความรู้ที่มีเหตุผลและทัศนคติทางศีลธรรม: “รสชาติ... ไม่มีอะไรมากไปกว่าความสามารถในการรู้สึกถึงความงาม เช่นเดียวกับเหตุผลคือความสามารถในการรับรู้ถึงความจริง ความรู้สึกที่สมบูรณ์ แท้จริง และศีลธรรม คือความสามารถในการรู้สึกดี"
ปัญหาเรื่องรสชาติได้รับการหยิบยกขึ้นมาในทฤษฎีสุนทรียภาพไปยังสถานที่ชั้นนำแห่งหนึ่งโดยเริ่มจากยุคฟื้นฟูศิลปวิทยาซึ่งเป็นภาพสะท้อนของปรากฏการณ์การทำให้ประสบการณ์ทางจิตวิญญาณของแต่ละบุคคลเป็นรายบุคคล ในด้านการพัฒนาทางศิลปะ เธอเคลื่อนตัวออกห่างจากหลักการ และในการประเมินด้านสุนทรียศาสตร์ เธอเริ่มถอยห่างจากแนวความคิดที่เป็นที่ยอมรับเกี่ยวกับความหมายของความสมบูรณ์แบบ ด้วยการผลักดันขอบเขตของสิ่งที่กำหนดไว้ในประสบการณ์ด้านสุนทรียศาสตร์ ผู้ถือรสนิยมจะยืนยันแง่มุมใหม่ของคุณค่าของปรากฏการณ์หรือเสนอวิสัยทัศน์ใหม่ให้กับสิ่งเหล่านั้น ดังนั้นแนวคิดในยุคกลางเกี่ยวกับความงามทางร่างกายในฐานะที่เป็นบาปจึงถูกแทนที่ด้วยการยืนยันถึงความงามทางร่างกาย เพลงสรรเสริญจึงถูกขับร้องเพื่อความกลมกลืนของหลักการทางร่างกายและจิตวิญญาณในมนุษย์ เราพบความคิดที่น่าสนใจเกี่ยวกับแก่นแท้ของรสนิยมในบทความของบุคคลสำคัญแห่งยุค L. Valla, M. Ficino, Pico de la Miran-dola, Leonardo da Vinci
ในศตวรรษที่ 17 แนวคิดของ "รสนิยม" เริ่มถูกนำมาใช้ในความหมายที่ชัดเจนโดยเฉพาะอย่างยิ่งต้องขอบคุณผลงานของนักปรัชญาชาวสเปน Gracian y Morales ("ฮีโร่", "ฉลาด" ฯลฯ ) ให้เราระลึกว่านักปรัชญาในศตวรรษที่ 17 - 18 ให้ความสนใจอย่างมากกับการพัฒนาทฤษฎีรสชาติ ในฝรั่งเศส (Battier, La Rochefoucauld, Tremblay, Rousseau, Helvetius, Voltaire) ในอังกฤษ (Shaftesbury, Hutcheson, Burke, Hume, Mandeville) ในเยอรมนี (Winckelmann, Lessing, Herder, Sulzer, Kant, Schiller) ความสนใจของนักวิจัยมุ่งเน้นไปที่คำถามเกี่ยวกับธรรมชาติของรสชาติ: มันเป็นเหตุผลหรือไม่มีเหตุผล ขึ้นอยู่กับเหตุผลหรือความรู้สึก เป็นรสชาติที่ได้รับจากการศึกษา หรือเป็นความสามารถโดยกำเนิด La Rochefoucauld ทำให้เกิดคำถามเกี่ยวกับการกำหนดรสนิยมส่วนบุคคล (บทความ "Maxims") วอลแตร์ในงานของเขา "เพลิดเพลิน" อธิบายลักษณะของปรากฏการณ์นี้เป็นการตอบสนองทางประสาทสัมผัสต่อความดีและความชั่วโดยพิจารณาจากความสามารถของสติปัญญาในการแยกแยะคุณสมบัติวัตถุประสงค์ของโลกวัตถุประสงค์ วอลแตร์ระบุถึงการปรับเปลี่ยนรสนิยมทางสุนทรีย์เช่นเดียวกับรสนิยมทางศิลปะ นักปรัชญาให้คำจำกัดความแนวคิดเรื่อง "รสชาติ" ในระบบความรู้เกี่ยวกับสุนทรียศาสตร์ว่าเป็น "อุปมา" บางอย่างที่ออกแบบมาเพื่อแสดงถึงความอ่อนไหวต่อความสวยงามและความน่าเกลียดในศิลปะ ขึ้นอยู่กับความสามารถนี้ เขาแบ่งรสนิยมออกเป็นดี เลว และบิดเบี้ยว “รสนิยมทางศิลปะนั้นบิดเบี้ยว” วอลแตร์เขียน “แสดงออกมาด้วยความรักต่อวิชาที่ทำลายล้างจิตใจที่มีการศึกษา ชอบล้อเลียนมากกว่าสิ่งสูงส่ง เสแสร้ง และงดงามเหนือความงามที่เรียบง่ายและเป็นธรรมชาติ นี่เป็นโรคของจิตวิญญาณ ( ตัวเอียงโดย V.M.) 11 คุณลักษณะนี้ฟังดูมีความเกี่ยวข้องมากแม้ในปัจจุบันในเงื่อนไขของการอยู่ใต้บังคับบัญชาของวัฒนธรรมทางศิลปะตามความต้องการของตลาด การแพร่กระจายของรสนิยมที่ไม่ดีเป็นอันตรายต่อการพัฒนาจิตวิญญาณโดยรวมของแต่ละบุคคล
วอลแตร์เน้นย้ำว่ารสนิยมทางศิลปะเป็นผลมาจากการศึกษาที่ยาวนานและรอบคอบ มนุษย์ต้องค่อยๆ เรียนรู้ที่จะฟังและมองโลกธรรมชาติและเรียนรู้คุณค่าทางศิลปะ นิสัยและการไตร่ตรองทำให้เธอสามารถรู้สึกถึงความสุขได้ในทันทีโดยมองเห็นบางสิ่งที่เธอไม่สามารถเข้าถึงได้ก่อนหน้านี้ นักปรัชญาเน้นย้ำถึงปรากฏการณ์ของการทำให้รสนิยมเป็นปัจเจกบุคคลไม่เพียงแต่ในระดับบุคคลเท่านั้น แต่ยังรวมถึงระดับของประเทศโดยรวมด้วย: “รสนิยมได้รับการปลูกฝังอย่างช้าๆ ในประเทศหนึ่งๆ เพราะมันค่อยๆ รับรู้ถึงจิตวิญญาณของศิลปินที่เก่งที่สุด ”
วอลแตร์ยังให้การตีความปัญหาหนึ่งของทฤษฎีรสชาติในปัจจุบัน: ใคร ๆ ก็สามารถโต้แย้งเกี่ยวกับรสนิยมได้หรือไม่? นักปรัชญาแยกแยะอย่างชัดเจนว่ารสชาติเป็นคุณสมบัติทางสรีรวิทยาของร่างกายและรสชาติที่สวยงาม แน่นอนว่า ไม่มีการโต้เถียงเรื่องรสนิยมเมื่อพูดถึงความพึงพอใจที่เกี่ยวข้องกับความสุขทางกาย สิ่งใดที่เป็นที่พอใจของคนคนหนึ่งอาจไม่เป็นที่พอใจของอีกคนหนึ่งก็ได้ อย่างไรก็ตาม สิ่งนี้ใช้ไม่ได้กับงานศิลปะ “เนื่องจากมีความงามที่แท้จริงในงานศิลปะ รสนิยมที่ดีจึงทำให้แตกต่าง และรสชาติที่ไม่ดีซึ่งไม่รับรู้ และข้อบกพร่องของจิตใจ - ที่มาของรสชาติที่บูด - จะต้องได้รับการแก้ไข” ให้เราเน้นหลายประการ ประเด็นในความคิดเห็นที่แสดงออกซึ่งเกี่ยวข้องกับทฤษฎีสุนทรียศาสตร์และการปฏิบัติของการศึกษาเกี่ยวกับสุนทรียศาสตร์ ประการแรก แหล่งที่มาของการก่อตัวของรสนิยมที่พัฒนาแล้วคือความงาม แหล่งที่มาของความงามตามวัตถุประสงค์คือศิลปะซึ่งหมายความว่าเป็น "ปัจจัยที่กระตือรือร้นใน การก่อตัวของรสชาติที่พัฒนาแล้ว ประการที่สอง ความงามซึ่งมีอยู่อย่างเป็นกลางในงานศิลปะที่สมบูรณ์แบบ จำเป็นต้องมีการพัฒนาโครงสร้างทางประสาทสัมผัสและสติปัญญาเพื่อที่จะเปิดเผยคุณสมบัติของมันต่อผู้ถูกทดลอง ประการที่สาม การเข้าสู่โลกแห่งความงามเป็นไปได้เฉพาะภายใต้เงื่อนไขของการโต้ตอบที่ประสานกันของโครงสร้างทางจิตวิญญาณของวัตถุ: ความสามารถในการรับรู้ทางประสาทสัมผัสและกิจกรรมของจิตใจ ซึ่งเผยให้เห็นคุณภาพของเรื่องที่น่ากังวล นี่คือความสอดคล้องของเรื่องกับแนวคิดเรื่องความสมควร (พลังภายใน) และความสมบูรณ์แบบของการสำแดงออกมาในการทำงานโดยรวมทางจิตวิญญาณ
ทฤษฎีสุนทรียศาสตร์ทำให้ระดับรสนิยมแตกต่างออกไป ดังนั้น Helvetius (บทความเรื่อง “On the Mind”) จึงแบ่งสิ่งเหล่านี้ออกเป็นสองประเภท: “รสชาติแห่งนิสัย” และ “รสชาติที่ใส่ใจ” ตามการจำแนกประเภทที่แพร่หลายในปัจจุบัน รสชาติมีจำกัดและพัฒนา สำหรับทฤษฎีรสชาติและการฝึกปฏิบัติเพื่อให้ความรู้เกี่ยวกับรสชาติที่พัฒนาแล้ว เหตุผลของ Helvetius เกี่ยวกับความแตกต่างระหว่างรสชาติทั้งสองระดับนี้เป็นสิ่งสำคัญ นักปรัชญามองเห็นความแตกต่างในแนวคิดเกี่ยวกับแก่นแท้ของความงาม แม้ว่าทั้งสองประเภทจะขึ้นอยู่กับประสบการณ์ด้านสุนทรียภาพก็ตาม ประการแรก -“ รสชาติของ zvichka11 - โดดเด่นด้วยทักษะบางอย่างในการประเมินปรากฏการณ์การตัดสินรสนิยมของนักเลงประเภทนี้นั้นโดดเด่นด้วยความจริงที่ว่าพวกเขาถือว่ามีคุณค่าเฉพาะสิ่งที่ได้กำหนดไว้ในประสบการณ์ของพวกเขาแล้ว ไม่มี ลิ้มรสพวกเขาทันทีที่ไม่มีวัตถุสำหรับการเปรียบเทียบ "- เฮลเวเทียสเขียน ประเภทที่สอง - "รสชาติที่มีสติ" - ขึ้นอยู่กับความรู้เชิงลึกในเรื่องของการประเมินและประสบการณ์ทางจิตวิญญาณที่เกิดจากวัฒนธรรม ผู้ถือสามารถชื่นชม ปรากฏการณ์ทางศิลปะใหม่และการประเมินจะเป็นไปตามวัตถุประสงค์การก่อตัวของรสนิยมประเภทนี้เกิดขึ้นได้จากการศึกษางานศิลปะและแนวคิดทางวิทยาศาสตร์ในระยะยาวเผยให้เห็นความรู้เกี่ยวกับความสวยงามอย่างแท้จริง
หน้าที่ด้านการศึกษาในการสร้างรสนิยมที่พัฒนาแล้วของสังคมในศตวรรษที่ 18-19 ดำเนินการโดยการวิจารณ์วรรณกรรมและศิลปะซึ่งในขณะนั้นถือเป็นกิจกรรมสุนทรียศาสตร์ประเภทพิเศษ เธอมีบทบาทเชิงบวกอย่างมากในการก่อตัวของวัฒนธรรมยุโรป (รวมถึงยุโรปตะวันออก) โดยการปลุกความสนใจในสมบัติที่สูงส่งและสวยงามที่สุดของวัฒนธรรมระดับชาติและระดับโลก ชี้แนะสาธารณชนให้หันไปหาตัวอย่างดังกล่าว และสร้างเกณฑ์สำหรับการรับรู้เชิงสุนทรีย์และการตัดสินรสชาติ ในการวิเคราะห์เชิงสุนทรีย์ของพวกเขา น่าเสียดายที่ในวัฒนธรรมหลังสมัยใหม่ประเพณีนี้สูญหายไป ดังนั้นรสนิยมของประชาชนจึงถูกทำลาย
สุนทรียศาสตร์แห่งความรู้สึกนิยมแบบอังกฤษสำรวจโครงสร้างที่ซับซ้อนของรสนิยมเชิงสุนทรียภาพ ตามคำกล่าวของ E. Burke รสชาติถูกสร้างขึ้นจาก "ความพึงพอใจเบื้องต้นของประสาทสัมผัสจากการรับรู้ปรากฏการณ์ ความหวานรองจากจินตนาการ และข้อสรุปของจิตใจเกี่ยวกับความสัมพันธ์ต่างๆ ระหว่างสิ่งเหล่านั้น ตลอดจนเกี่ยวกับกิเลสตัณหา ศีลธรรม และการกระทำของมนุษย์" ดังนั้น รสชาติจึงไม่ใช่การสำแดงออกของราคะที่เกิดขึ้นในทันทีเช่นนี้ ไม่ใช่ขอบเขตของความไร้เหตุผลล้วนๆ แต่ก็ไม่ใช่ขอบเขตของแนวคิดล้วนๆ ด้วย รสชาติเป็นปฏิสัมพันธ์อินทรีย์ของความสุขของประสาทสัมผัส ความสุขของจินตนาการ และข้อสรุปของจิตใจ นอกจากนี้ ยังตั้งข้อสังเกตด้วยว่าปฏิสัมพันธ์ที่มีชื่อของโครงสร้างทางปัญญาและประสาทสัมผัสนั้นเป็นเรื่องปกติสำหรับทัศนคติเชิงสุนทรีย์ทุกประเภท และรสนิยมสามารถปรับปรุงได้อย่างต่อเนื่องโดยการขยายประสบการณ์ของการรับรู้ เพิ่มคุณภาพของวัตถุให้ลึกซึ้งยิ่งขึ้น และฝึกฝนอย่างต่อเนื่องในการรับรู้เกี่ยวกับสุนทรียศาสตร์ ให้เราใส่ใจกับความครอบคลุมของปรากฏการณ์ตามที่นักปรัชญาได้เปิดเผยไว้ รสชาติถือเป็นความสามารถทางปัญญาและเป็นผลมาจากการเลือกวัตถุอย่างมีสติและการสร้างทัศนคติที่เป็นรายบุคคลต่อสิ่งเหล่านั้น สิ่งสำคัญคือขอบเขตของประสบการณ์ด้านสุนทรียภาพที่เป็นปัจเจกบุคคล ซึ่งตัดสินตัวเองด้วยการตัดสินรสนิยม ไม่ได้กำหนดปรากฏการณ์ใด ๆ แต่เป็นปรากฏการณ์ทางศิลปะที่สมบูรณ์แบบ นั่นคือ ผู้ถือครองคุณค่าอันเป็นสากลในเนื้อหา สิ่งหลังมีความสำคัญในการสร้างทฤษฎีทางวิทยาศาสตร์เกี่ยวกับการศึกษาด้านสุนทรียภาพและการศึกษาด้วยตนเองของแต่ละบุคคล ทฤษฎีสุนทรียศาสตร์คลาสสิกมองว่าบุคคลเป็นหัวข้อที่กระตือรือร้นของทัศนคติเชิงสุนทรียศาสตร์และขจัดแนวคิดเรื่องความสัมพันธ์ออกจากกระบวนการศึกษาเกี่ยวกับสุนทรียภาพทั้งในเนื้อหาของค่านิยมและในวัตถุประสงค์
ให้เรากลับไปสู่สุนทรียศาสตร์คลาสสิกของเยอรมันโดยมุ่งเน้นไปที่ทฤษฎีรสนิยมในสุนทรียศาสตร์ของคานท์ที่พัฒนาขึ้นในผลงาน: "การสังเกตความรู้สึกของความสวยงามและประเสริฐ" (1764), "บทวิจารณ์พลังแห่งการพิพากษา" (1790) “มานุษยวิทยาในความสัมพันธ์เชิงปฏิบัติ” (1798) ในการพัฒนาทฤษฎีแห่งรสนิยม Kant อาศัยแนวคิดของ E. Burke, D. Hume และนักปรัชญานักกระตุ้นความรู้สึกชาวอังกฤษคนอื่น ๆ ผลงานของ Kant ให้เหตุผลสำหรับธรรมชาติของรสนิยมนิรนัย ยืนยันแนวคิดความเป็นสากลของการตัดสินรสชาติและตรวจสอบประเด็นหลักสี่ประการของรสชาติที่เกี่ยวข้องกับ "เกมความสามารถทางปัญญา" ประเด็นแรกประกอบด้วยแนวคิดของ "การไม่ครอบงำจิตใจ 11 การตัดสินรสชาติ มันพิสูจน์ว่ารสชาติมีความเกี่ยวข้องกับความพึงพอใจทางสุนทรีย์ซึ่งกำหนดโดยคุณสมบัติของวัตถุแห่งความเฉยเมย ช่วงเวลาที่สองและสี่ยืนยันถึงความสวยงามเช่นนี้ ซึ่งในการตัดสินรสชาตินั้นรับรู้ได้โดยปราศจากแนวคิด เนื่องจากมันเป็น "วัตถุแห่งความเพลิดเพลินที่จำเป็น" นั่นคือพื้นฐานของรสนิยมคือความรู้สึกถึงความงาม ศิลปะที่แสดงถึงความงามที่ตระการตาได้รับการเปิดเผยโดย I. Kant ในฐานะแหล่งที่มาของวิธีการรับรู้แบบพิเศษ - การรับรู้ในภาพที่ให้ความสุขทางจิตวิญญาณอย่างมหาศาลในรูปแบบที่สมบูรณ์แบบ การรับรู้ที่มีพื้นฐานมาจากความหวานนั้นไม่มีใครสังเกตเห็น: ความสามารถทางปัญญาดูเหมือนจะเล่นมากกว่าทำงาน ด้านที่สามของการตัดสินรสชาติยืนยันคุณค่าที่แท้จริง - "ความมุ่งหมายที่ไม่มีเป้าหมาย" เนื่องจากเป้าหมายของการตัดสินรสชาติทางสุนทรีย์นั้นเป็นเป้าหมายสำหรับประสาทสัมผัสเนื่องจากความมีชีวิตชีวาภายในที่มีจุดประสงค์และสมบูรณ์แบบ คานท์เปรียบเทียบงานศิลปะโดยคำนึงถึงจุดมุ่งหมายภายในกับธรรมชาติในจุดประสงค์ของรูปแบบโดยธรรมชาติ โดยคำนึงถึงความแตกต่างระหว่างสิ่งเหล่านั้น นักปรัชญาตั้งข้อสังเกตถึงความสมบูรณ์แบบภายในของงานศิลปะเมื่อทำหน้าที่เป็นหลักการก่อตัวทางจิตวิญญาณ รวบรวมโครงสร้างทางจิตวิญญาณของมนุษย์ให้เป็นความสมบูรณ์ ผสมผสานการเชื่อมโยงกันของสติปัญญาและความรู้สึกในการตัดสินรสนิยม นักปรัชญาตีความรสชาติว่าเป็นความสามารถในการประเมิน (ปรากฏการณ์ทางธรรมชาติ ปรากฏการณ์ทางศิลปะ) แม้กระทั่งปล่อยให้ "ความรู้สึก... ได้รับการถ่ายทอดไปยังคนอื่นๆ" กล่าวอีกนัยหนึ่ง: การโน้มน้าวใจทางศิลปะของงานสามารถปลุกเร้าและสร้างความรู้สึกและสติปัญญาได้กระตุ้นให้เราตัดสินรสนิยมทางสุนทรีย์ที่เพียงพอกับคุณสมบัติของงาน
คานท์มองว่าวิภาษวิธีของแต่ละบุคคลและส่วนรวมเป็นปัญหาสำคัญในการตัดสินรสนิยมทางสุนทรีย์ หากการตัดสินของแต่ละบุคคลรวมถึงหลักการของความเป็นสากลด้วย หลักการนี้ก็จะต้องมีอยู่ในความรู้สึกเชิงสุนทรีย์ในตัวของมันเอง ความรู้สึกพึงพอใจเกิดจากความได้เปรียบที่เป็นสากล ซึ่งนิยามโดยอัตวิสัยว่าเป็นหลักการเบื้องต้นของจิตสำนึก และโดยแท้จริงแล้วมันปรากฏเป็น "รูปแบบบริสุทธิ์" ของวัตถุ คานท์ชี้แจงเส้นทางสู่จิตสำนึกที่ได้รับประสบการณ์จากสากลโดยเสนอแนวคิดเรื่อง "จิตสำนึกสามประการในชีวิตประจำวัน" พวกเขาสามารถช่วยอธิบาย "การวิจารณ์รสชาติ" ได้ กล่าวคือ 1) มีวิจารณญาณของคุณเอง; 2) วางจิตใจให้อยู่ในตำแหน่งของคนอื่น 3) คิดให้สอดคล้องกับตัวเองเสมอ
นักปรัชญาชี้แจงแนวคิดว่าข้อแรกหมายถึงจุดสูงสุดของวิธีคิดที่ปราศจากอคติ ประการที่สองคือวิธีคิดกว้างๆ คือ ความสามารถในการตัดสินของตนเองให้ก้าวไปไกลกว่ามุมมองทั่วไป (ซึ่งจะพบได้จากการแบ่งปันความคิดเห็นของผู้อื่นเท่านั้น) ในที่สุด คติพจน์ที่สาม - วิธีคิดตามลำดับ - เกิดขึ้นได้จากการผสมผสานระหว่างหลักการที่หนึ่งและที่สองเท่านั้น และการโต้ตอบระหว่างสิ่งเหล่านั้นจนกลายเป็นทักษะ หลักการนี้บรรลุผลได้มากที่สุด คติพจน์ทั้งสามที่มีชื่อตามที่คานท์กล่าวไว้ ครอบคลุมขอบเขตทั้งหมดของสติปัญญา เนื่องจากประการแรกคือขอบเขตสูงสุดของความเข้าใจ ขอบเขตที่สองของความสามารถในการตัดสิน และประการที่สามของจิตใจ การเชื่อมโยงวิภาษวิธีของพวกเขาถูกสร้างขึ้นเช่นนี้: จินตนาการในอิสรภาพของมันปลุกการทำงานของจิตใจซึ่งโดยไม่ต้องอาศัยการไกล่เกลี่ยแนวคิดก็ปล่อยให้ความถูกต้องเป็นการเล่นของจินตนาการ: สิ่งที่ผู้อื่นนำเสนอนั้นไม่ใช่ความคิด แต่“ เป็นความรู้สึกภายในของสภาวะที่มีจุดมุ่งหมายของจิตวิญญาณ (ตัวเอียงของผู้เขียน - V.M. )” คานท์ถือว่าความรู้สึกและการตัดสินของรสนิยมเป็นความต้องการที่กำหนดโดยธรรมชาติของมนุษย์ - สิ่งมีชีวิตที่ถูกลิขิตให้มีชีวิตอยู่ในสังคมและด้วยเหตุนี้ รู้สึกถึงความจำเป็นในการสื่อสาร
ในสุนทรียศาสตร์ของคานท์ ความคิดเกี่ยวกับธรรมชาติที่ไม่สนใจในทางปฏิบัติของการตัดสินเชิงสุนทรีย์แห่งรสชาตินั้นได้รับการติดตามอย่างต่อเนื่อง การตัดสินรสนิยมโดยสนใจนั้นขึ้นอยู่กับสมมติฐาน ความสนใจในสิ่งที่ดีทางศีลธรรม และแนวโน้มของวิธีคิดที่ดีทางศีลธรรม การตัดสินรสนิยมเกี่ยวกับธรรมชาติและศิลปะเป็นวิธีการหนึ่งที่แสดงถึง “ความสะดวกโดยไม่มีจุดมุ่งหมาย” เพราะจำเป็นต้องสร้างวัฒนธรรมความสามารถของจิตวิญญาณในการสื่อสารระหว่างผู้คน มุมมองสากลของความสุขกำหนดว่าความสุขนี้ไม่ได้ขึ้นอยู่กับความรู้สึกเท่านั้น แต่ยังเป็นความสุขของการไตร่ตรอง นั่นคือ มันเกี่ยวข้องกับความสามารถในการสะท้อนกลับของการตัดสิน
ประวัติความเป็นมาของการก่อตัวของทฤษฎีรสชาติได้รับการวิเคราะห์โดยเห็นได้จากความเชื่อมโยงกับความจริงที่ค้นพบโดยวิทยาศาสตร์ความสามารถในการรับรู้ถึงอาการทางประสาทสัมผัสของความสมบูรณ์แบบในปรากฏการณ์ทางธรรมชาติและปรากฏการณ์ทางศิลปะอย่างเพียงพอความสามารถในการสะท้อนปรากฏการณ์การรับรู้และของตัวเอง ประสบการณ์ในคุณสมบัติของพวกเขาและในที่สุดความสามารถในการถ่ายทอดในการตัดสินความรู้สึกของคนที่เกิดจากวัตถุ ความแน่นอนเชิงคุณภาพของการตัดสินรสชาติที่พัฒนาโดยทฤษฎีสุนทรียภาพนั้นมีลักษณะเฉพาะ - นี่คือความจริง (ตัวอย่างที่ต้องการ, แบบจำลองในอุดมคติ) ในการฝึกฝนการตัดสินคุณค่านั้นแสดงออกว่าเป็นรสนิยมที่ได้รับการพัฒนาหรือสุนทรียภาพ ผู้ถือมันเป็นบุคคลที่มีประสบการณ์ทางจิตวิญญาณมากมาย มีความสามารถไม่เพียงแต่ตัดสินอย่างเป็นกลางเกี่ยวกับคุณค่าเท่านั้น แต่ยังสร้างสิ่งเหล่านั้นด้วย มันเป็นลักษณะของสัดส่วนในการแสดงออกการปรากฏตัวของเกณฑ์ในการตัดสินเชิงสุนทรียศาสตร์และในความสัมพันธ์กับโลก (ทัศนคติต่อผู้อื่นต่อคุณค่าทางศีลธรรมและศิลปะของสังคมและมนุษยชาติ ฯลฯ ) ประสบการณ์ของเธอเกี่ยวกับปรากฏการณ์และการตัดสินเกี่ยวกับสิ่งเหล่านั้นนั้นมีเอกลักษณ์เฉพาะตัวโดยมีความสอดคล้องกับความเป็นสากลของเนื้อหาของการตัดสิน (หลักฐานการครอบครองความจริงเกี่ยวกับสิ่งต่าง ๆ )
นอกจากรสชาติที่พัฒนาแล้ว ทฤษฎีสุนทรียศาสตร์สมัยใหม่ยังระบุถึงรสชาติประเภทที่ไม่ดีหรือเสียไป (บิดเบี้ยว) ผู้ที่มีรสนิยมเช่นนั้นย่อมได้รับความเพลิดเพลินจากการใคร่ครวญถึงสิ่งที่น่าเกลียดและไม่แยแสกับความงาม อันตรายของแนวความคิดอันทรงคุณค่าดังกล่าวคือการที่ผู้ถือเอาความคิดเชิงลบแพร่กระจายการแสดงออกของมนุษย์ ในรูปแบบศิลปะและการปฏิบัติ โดยปลูกฝังให้เป็นบรรทัดฐานในการทำความเข้าใจธรรมชาติของสิ่งที่มีค่า ยิ่งไปกว่านั้น วิธียืนยันค่าหลอกนั้นมีลักษณะก้าวร้าวซึ่งสอดคล้องกับหัวข้อของการยืนยัน อันตรายของเนื้อหาของแนวคิดเชิงคุณค่าและการตัดสินคุณค่าดังกล่าวก็คือ เนื้อหาเหล่านี้สามารถทำลายรสนิยมของสาธารณชน แพร่กระจายความสนใจในสิ่งที่น่าเกลียดและผิดศีลธรรมภายใต้สโลแกน "ทันสมัย" และ "ดั้งเดิม"
เกี่ยวกับลักษณะเชิงปริมาณรสชาติที่ยังไม่พัฒนา ("แคบ") มีความโดดเด่นถัดจากรสชาติที่พัฒนาแล้ว ผู้ถือครองคือผู้ที่มีวัฒนธรรมในระดับต่ำและมีประสบการณ์ในการสื่อสารกับคุณค่าทางศิลปะอย่างจำกัด พวกเขาขาดเกณฑ์ความดีและความชั่ว สวยงามและน่าเกลียด พวกเขาโดดเด่นด้วยการขาดเหตุผลในการประเมินและการทำอะไรไม่ถูกในการตัดสินเกี่ยวกับคุณภาพของวัตถุ พวกเขาไม่สามารถอธิบายตัวเองได้ว่าทำไมและอะไรดึงดูดพวกเขาให้สนใจเรื่องนี้หรือเรื่องนั้น การประเมินไม่รวมถึงเนื้อหาทั่วไปของแนวคิดเชิงคุณค่า การตัดสินเหล่านี้ขึ้นอยู่กับประสบการณ์เชิงประจักษ์และดังนั้นจึงเป็นไปตามอำเภอใจ เป็นที่น่าสังเกตว่าในการสื่อสารพวกเขาเลือกสิ่งที่เข้าใจได้และน่าพึงพอใจสำหรับตัวเอง ดังนั้นพวกเขาจึงอยู่ในวงจรของประสบการณ์ที่จำกัด สินค้ามาตรฐานจะทำให้รสนิยมเป็นกลางและทำให้เกิดความซ้ำซากจำเจในการตัดสินเกี่ยวกับคุณภาพ
โปรดทราบว่าสามารถให้ความรู้เกี่ยวกับรสชาติที่ยังไม่พัฒนาและภายใต้การสื่อสารอย่างเป็นระบบด้วยคุณค่าทางสุนทรีย์และผู้ให้บริการของรสชาติที่พัฒนาแล้วสามารถลึกซึ้งและปรับปรุงได้ การก่อตัวของรสนิยมทางสุนทรีย์ที่พัฒนาแล้วเป็นเป้าหมายของกระบวนการสอน เนื่องจากเด็กมักจะมีรสนิยมที่ยังไม่พัฒนาเนื่องจากมีประสบการณ์น้อยในการสื่อสารกับคุณค่าทางศิลปะ อย่างไรก็ตาม เป็นที่น่าสังเกตว่าในระดับสัญชาตญาณ พวกเขาสามารถเข้าใจปรากฏการณ์ที่แสดงออกทางสุนทรียศาสตร์ได้แม่นยำกว่าผู้ใหญ่ แม้ว่าพวกเขาจะไม่สามารถประเมินตัวเลือกของตนเองและกำหนดเกณฑ์การประเมินได้ก็ตาม
ให้เรามุ่งเน้นไปที่คุณค่าทางสังคมของรสนิยมทางสุนทรีย์ที่พัฒนาแล้ว โดยคำนึงถึงความสามารถในการพัฒนาทั้งในด้านความสัมพันธ์กับปัจเจกบุคคลและในความสัมพันธ์กับสังคมในฐานะจิตวิญญาณทั้งหมด โดยปกติแล้วระดับรสชาติที่สำคัญจะถูกกำหนดโดย "จิตวิญญาณแห่งกาลเวลา" ความสนใจในความจริงหรือในทางกลับกันการเพิกเฉยนั้นสะท้อนให้เห็นในลักษณะของการตอบสนองต่อปรากฏการณ์ของผู้คนจำนวนมาก (การพูดคุยเกี่ยวกับรสนิยมของยุคสมัยประเทศชนชั้น ฯลฯ เป็นเรื่องถูกต้องตามกฎหมาย) คานท์ให้เหตุผลว่ารสชาติที่ดีจะปรากฏออกมาเฉพาะในช่วงเวลาที่ดีต่อสุขภาพเท่านั้น ไม่ใช่แค่รสชาติที่ประณีตเท่านั้น รสนิยมที่พัฒนาแล้วทำให้ความรู้สึกเป็นสากล นำพวกเขาไปสู่ความเข้าใจในความสัมพันธ์สากลในประสบการณ์ทางจิตวิญญาณ และในขณะเดียวกันก็ทำให้พวกเขาเป็นรายบุคคล การแสดงออกถึงรสนิยมที่เป็นรายบุคคลนั้นมีคุณค่าเพราะมันแสดงให้เห็นถึงความแตกต่างของคุณสมบัติของสิ่งที่เป็นกังวลและช่วยให้คุณถ่ายทอดความรู้สึกที่บุคคลนั้นได้รับ ด้วยเหตุนี้ จึงมีการสร้างพื้นที่สำหรับการสร้างร่วมในการโต้ตอบระหว่างการประเมินและประสบการณ์ ดังนั้นรสนิยมทางสุนทรีย์จึงเป็นวิธีการที่มีประสิทธิภาพในการรวมจิตวิญญาณของผู้คน
โปรดทราบว่าหน้าที่ด้านการสร้างจิตวิญญาณนั้นดำเนินการโดยรสนิยมที่ "ดี" ที่พัฒนาแล้วเท่านั้น โดยอาศัยประสบการณ์อันยาวนานในการสื่อสารด้วยคุณค่าทางสุนทรีย์ ผลกระทบทางการศึกษาที่มีประสิทธิผลมากที่สุดคือวัฒนธรรมทางศิลปะ ประการแรกคือมรดกทางศิลปะคลาสสิก ซึ่งเกิดขึ้นจากการเป็นตัวตนที่สมบูรณ์แบบของประสบการณ์ด้านสุนทรียภาพระดับสูง ความสมบูรณ์ของเนื้อหาที่มีคุณค่าและความสมบูรณ์แบบของภาพทางศิลปะของการดำรงอยู่ของมันกำหนดประสิทธิภาพของความเป็นไปได้ในการก่อสร้างในขอบเขตแห่งจิตวิญญาณ ให้เรากลับมาสนใจความคิดเห็นของ อ.คานต์ อีกครั้ง รสนิยมที่ดี เน้นย้ำถึงสิ่งที่สอดคล้องกับ “แนวคิดของสิ่งของ” และความหมายของการเลือกคือความสามารถในการเลือกสิ่งที่ทุกคนชอบ กล่าวคือ การเลือกด้วย ความรู้สึกที่จะเข้าสังคม ขอให้เราใส่ใจกับความจริงที่ว่าวัตถุแห่งความรู้สึกและคุณค่าของการตัดสินรสชาติไม่ใช่วัตถุเช่นนั้น (ความเป็นจริงทางวัตถุ) แต่เป็นคุณค่าทางจิตวิญญาณของความหมาย ยิ่งหัวข้อของกิจกรรมการประเมินเจาะลึกเข้าไปในฐานะหัวข้อของการตัดสินที่มีประสบการณ์ การตัดสินรสชาติก็จะสมบูรณ์และลึกซึ้งมากขึ้นเท่านั้น และประสบการณ์ของมันก็เผยออกมาในการตัดสินในลักษณะที่มีเอกลักษณ์เฉพาะตัวและสร้างสรรค์ในธรรมชาติ ผู้ถือรสนิยมที่พัฒนาแล้วสามารถระบุได้ว่าอะไรในคำพูดของอิมมานูเอล คานท์ “เป็นที่ชื่นชอบมาเป็นเวลานาน”
บทความในหัวข้อ